
American Journal of Botany87(5):601–607.2000.
INVITED SPECIAL PAPER
A PICAL CONTROL OF BRANCH GROWTH AND ANGLE IN
WOODY PLANTS1
B RAYTON F.W ILSON
Department of Natural Resources Conservation,University of Massachusetts,Amherst,Massachusetts01003USA
Apical control is the inhibition of a lateral branch growth by shoots above it(distal shoots).If the distal shoots are cut off to remove apical control,the lateral branch can grow larger and may bend upwards.Apical control starts when new lateral buds grow after passing through a period of dormancy.Buds initially break and produce leaves,then apical control is exerted and the lower(proximal)laterals stop growing.Apical control also inhibits growth of large,old branches.
Gravimorphism and restricted water and nutrient transport can inhibit branch growth,but they are not primary mechanisms of apical control.Apical control may reduce branch photosynthesis.Under apical control allocation of branch-produced assimilate to the stem is relatively high,so low assimilates in the branch may limit branch growth even though hormone levels are adequate for growth.Hormones appear to be involved in apical control,but it is not known how.One role of hormones may be to maintain the strength of the stem sink for branch-produced assimilate.Upward bending of a woody branch after release from apical control requires both new wood production and production of wood cells that can generate an upward bending moment.Apical control inhibits radial growth of branches and,in some species,may regulate the production of wood with an upward bending moment.
Key words:allocation;apical control;branch angle;branch growth;growth stress.
Woody plants have characteristic branching patterns (Fig.1).Lateral branches are usually smaller,and at a larger angle to vertical,than their parent shoots.Also, branches of the same age generally decrease in size down their parent shoot.As a result,many species have branch-ing patterns with long shoots at the upper(distal)end of each annual increment and short shoots at the lower (proximal)end.The short shoots produce leaves,but scarcely elongate.
These branching patterns are the result of apical con-trol.The parent and lateral shoots above a lateral branch inhibit the growth of the branch.This inhibition is called apical control.The inhibition can be demonstrated by de-capitating the shoots above the branch or by a phloem girdle just above the branch(Fig.1).After removing api-cal control the distal branch grows larger,and often grows more vertically,than if it had remained inhibited. When apical control is removed,a distal branch often replaces the injured terminal,reimposes apical control, and maintains the characteristic branching pattern.
There are different types of lateral branches in woody plants.Sylleptic lateral branches develop from lateral meristems without a period of dormancy while the ter-minal shoot is elongating;proleptic lateral branches de-velop from lateral buds after a period of dormancy(Halle′, Oldeman,and Tomlinson,1978).Brown,McAlpine,and Kormanik(1967)proposed the term‘‘apical control’’to describe the inhibition of growth of proleptic branches in contrast to the term‘‘apical dominance’’to describe bud formation though inhibition of growth of sylleptic branches.Thus,in temperate plants when a lateral meri-
1Manuscript received7July1998;revision accepted9July1999.
The author thanks the many researchers,students,and friends he has worked with over the years.This material is based upon work partially supported by Cooperative State Research,Extension,Education Ser-vice,U.S.Department of Agriculture,Massachusetts Agricultural Ex-periment Station,under project number McIntire-Stennis70.
stem is formed apical dominance determines whether the meristem initially forms a sylleptic branch or forms a bud.In the second year apical control regulates the amount of elongation and diameter growth of proleptic branches from the previously dormant buds.Some buds do not form proleptic branches after the initial period of dormancy.These buds may enter the bud bank and sub-sequently form epicormic branches,a third type of branch (Wilson and Kelty,1994).Epicormic branches generally grow rapidly,are often near-vertical,and are the source of reiteration in tropical trees(Halle′,Oldeman,and Tom-linson,1978).Epicormic branches may function as re-placement branches in species whose branches cannot bend up,such as the plagiotropic branches of Araucaria (House et al.,1998).In this paper I will restrict the term apical control to the regulation of the primary and sec-ondary growth of proleptic lateral branches,both young and old,that have grown from a bud following a single period of dormancy.
There are minor disagreements about terminology.
Cline(1997)de?nes apical dominance in herbaceous plants‘‘as the control exerted by the shoot apex over the outgrowth of the lateral buds.’’He proposes that the term apical control applies to the control exerted on lateral shoots after the bud has started elongating,even though the bud has not passed through a period of dormancy.
Europeans use the term‘‘acrotony,’’somewhat compa-rable to apical control,to indicate the dominant growth of the uppermost,distal,laterals after bud dormancy in woody plants(Champagnat,1978).We should not let dif-ferences in terminology obscure the probability that as the phenomena of branch growth change during devel-opment from lateral meristem formation to a large, woody,lateral branch,the mechanisms controlling growth also change.Mechanisms of apical dominance in herbaceous plants should not just be assumed to apply to apical control phenomena in all stages of branch growth in woody plants.
601
602[Vol.87
A MERICAN J OURNAL
OF
B
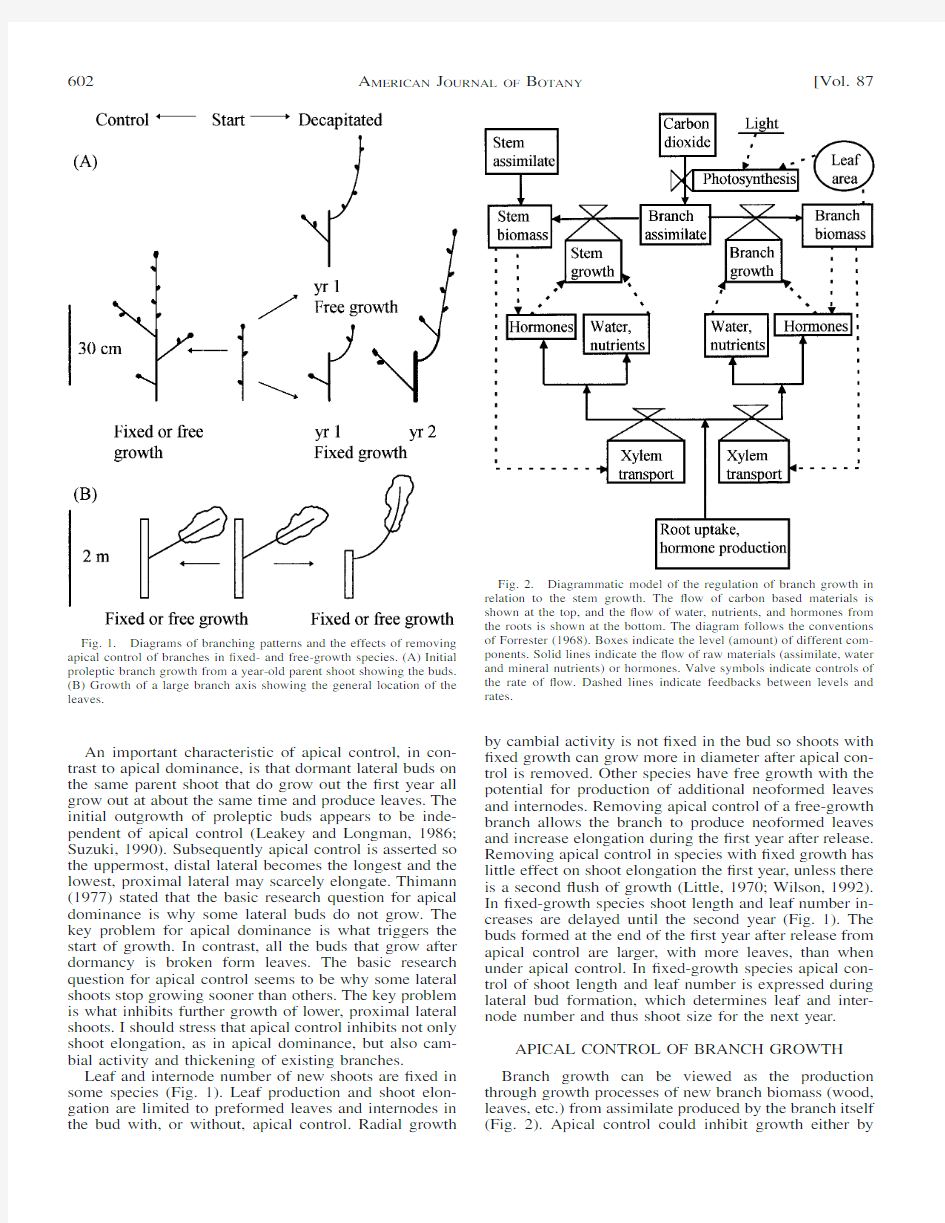
OTANY Fig.1.Diagrams of branching patterns and the effects of removing apical control of branches in ?xed-and free-growth species.(A)Initial proleptic branch growth from a year-old parent shoot showing the buds.(B)Growth of a large branch axis showing the general location of the
leaves.
Fig.2.Diagrammatic model of the regulation of branch growth in relation to the stem growth.The ?ow of carbon based materials is shown at the top,and the ?ow of water,nutrients,and hormones from the roots is shown at the bottom.The diagram follows the conventions of Forrester (1968).Boxes indicate the level (amount)of different com-ponents.Solid lines indicate the ?ow of raw materials (assimilate,water and mineral nutrients)or hormones.Valve symbols indicate controls of the rate of ?ow.Dashed lines indicate feedbacks between levels and rates.
An important characteristic of apical control,in con-trast to apical dominance,is that dormant lateral buds on the same parent shoot that do grow out the ?rst year all grow out at about the same time and produce leaves.The initial outgrowth of proleptic buds appears to be inde-pendent of apical control (Leakey and Longman,1986;Suzuki,1990).Subsequently apical control is asserted so the uppermost,distal lateral becomes the longest and the lowest,proximal lateral may scarcely elongate.Thimann (1977)stated that the basic research question for apical dominance is why some lateral buds do not grow.The key problem for apical dominance is what triggers the start of growth.In contrast,all the buds that grow after dormancy is broken form leaves.The basic research question for apical control seems to be why some lateral shoots stop growing sooner than others.The key problem is what inhibits further growth of lower,proximal lateral shoots.I should stress that apical control inhibits not only shoot elongation,as in apical dominance,but also cam-bial activity and thickening of existing branches.
Leaf and internode number of new shoots are ?xed in some species (Fig.1).Leaf production and shoot elon-gation are limited to preformed leaves and internodes in the bud with,or without,apical control.Radial growth
by cambial activity is not ?xed in the bud so shoots with ?xed growth can grow more in diameter after apical con-trol is removed.Other species have free growth with the potential for production of additional neoformed leaves and internodes.Removing apical control of a free-growth branch allows the branch to produce neoformed leaves and increase elongation during the ?rst year after release.Removing apical control in species with ?xed growth has little effect on shoot elongation the ?rst year,unless there is a second ?ush of growth (Little,1970;Wilson,1992).In ?xed-growth species shoot length and leaf number in-creases are delayed until the second year (Fig.1).The buds formed at the end of the ?rst year after release from apical control are larger,with more leaves,than when under apical control.In ?xed-growth species apical con-trol of shoot length and leaf number is expressed during lateral bud formation,which determines leaf and inter-node number and thus shoot size for the next year.
APICAL CONTROL OF BRANCH GROWTH Branch growth can be viewed as the production through growth processes of new branch biomass (wood,leaves,etc.)from assimilate produced by the branch itself (Fig.2).Apical control could inhibit growth either by
May2000]603
W ILSON—A PICAL CONTROL
affecting the growth process or by limiting the raw ma-terials needed for growth.Hormones,water stress,and nutrients can affect growth rates.The major raw material for growth is assimilate from photosynthesis,but water and mineral nutrients are also required.Therefore,apical control could reduce branch growth either by affecting hormones or water even with suf?cient assimilate,or by restricting the availability of assimilate even with ade-quate hormones.Water and some hormones move into the branch through the xylem so they are affected by xylem transport.Other hormones are produced by the branch itself.Production of branch assimilate is a func-tion of branch photosynthesis.The rate of photosynthesis is determined by a number of internal and external fac-tors.Once branch assimilate is produced it is then allo-cated to either the branch or the stem.
Presumably environmental effects act by modifying some component of the model in Fig.2,so I will not discuss them separately.Some environmental effects on apical control may re?ect the observation that slow-grow-ing plants have lower apical control.Apical control is reduced in conifer trees in the shade(O’Connell and Kel-ty,1994),or higher order,slower growing,branches (Remphrey and Davidson,1992),or in old slow-growing trees(Moorby and Wareing,1963).Therefore,any en-vironmental effect that reduced overall growth rates could indirectly reduce apical control.
Most research has been done on?rst-order lateral branches(using the botanical numbering system)arising from a vertical,central,parent shoot.I will call this?rst-order shoot the stem.A?rst-order branch off the stem may live for decades.The terminal of the stem,or of a branch,is the current annual elongation growth.There-fore,parent shoots and lateral branches of all orders all have terminals.
Hormones—The hormonal hypothesis for apical dom-inance was developed by Thimann and Skoog to explain apical dominance in herbaceous species(Thimann,1977). The original proposed mechanism was that auxin from the parent terminal directly inhibited lateral bud growth. Subsequently the mechanism was modi?ed to include the action of other hormones,particularly cytokinins(Thi-mann,1977).Strong support for the effect of auxin comes both from auxin replacement experiments where auxin applied to a decapitated terminal shoot inhibits growth of lateral buds and from experiments using TIBA (tri-iodo benzoic acid)to block auxin transport and re-move apical dominance(Cline,1997).
Auxin is transported polarly from shoot tip toward the roots.In Pinus sylvestris both the terminal shoot and branches contribute auxin to the stem,and the stem can also produce some of its own auxin(Sundberg and Ug-gla,1998).The polar nature of apical control parallels the polar transport of auxin.Generally the uppermost, distal shoots inhibit the lower,proximal shoots.Several experiments modify this polar https://www.doczj.com/doc/57354638.html,terals of Po-pulus deltoides seedlings were all the same length,rather than showing the‘‘normal’’decrease in length from tip to base,when the plants are grown under long days, warm temperatures,and high nutrients(Richards and Lar-son,1981).Changing the angle of rooted cuttings of Tri-plochiton scleroxylon eliminated,or even reversed,the pattern of lateral shoot lengths(Leakey and Longman, 1986).It seems reasonable that treatments that affect pat-terns of shoot length also affect polar transport,but the hypothesis has not been tested.
There are relatively few experiments on hormonal ac-tion in apical control.Auxin in high concentrations ap-plied to the decapitated or girdled stem of a woody plant can replace apical control and inhibit branch diameter growth,upward bending,and production of neoformed leaves in free growth species(Wilson and Archer,1983; Timell,1986,pp.1189–1193;House et al.,1998).Al-though TIBA apparently blocks auxin transport through woody shoots(Kennedy and Farrar,1965),I am not aware of experiments using TIBA or other auxin trans-port inhibitors to remove apical control over woody branches.Some experiments do not seem to support the auxin hypothesis for apical control.In Pinus strobus,a girdle just below a branch removes apical control even though the branch and the terminal shoot are still in direct connection and movement of hormones from terminal to lateral was,presumably,not affected(Mu¨nch,1938;Wil-son,1981).Therefore,although auxin probably is in-volved in apical control,it is not clear how.
There is good evidence that hormones in addition to auxin regulate the amount of lateral shoot growth in plants.High cytokinin to auxin ratios were associated with fast growth of laterals in Lupinus,a nonwoody plant, (Emery,Longnecker,and Atkins,1998).In Picea abies, a?xed-growth tree,there was a positive correlation be-tween cytokinin(zeatin riboside)content and lateral bud size during the critical period of bud formation that de-termines shoot length(Chen,Bollmark,and Eliasson, 1996).Gibberellins can overcome apical control in a number of conifer species and ethylene also appears to be involved(Timell,1986,pp.1197–1199).
Experimental applications of exogenous hormones clearly implicate hormones in both apical dominance and apical control.Measurements of endogenous hormone levels in relation to elongation and cambial activity in woody plants are often inconsistent with results from ex-ogenous treatments(Little and Pharis,1995).The?eld of hormonal control seems to be in a phase of reassessment. The next step may be,as suggested by Cline(1994),the use of transgenic plants with different endogenous hor-mone levels.Unfortunately for advancement in knowl-edge of apical control,current work on transgenic plants is primarily on herbaceous plants(Schmitz and Theres, 1999).
Transport of water and mineral nutrients—One hy-pothesis is that transport of water and nutrients to the lateral branch is restricted relative to transport to the ter-minal shoot and therefore branch growth is reduced rel-ative to the terminal.Borchert and Honda(1984)used this hypothesis to successfully simulate branching in a tree by assuming that growth was controlled by?ux dis-tribution among branches.Studies of hydraulic architec-ture support the hypothesis.In trees the hydraulic con-ductivity of branches is lower than the conductivity of the terminal,particularly at the junction between the ter-minal and the branch(Zimmermann,1978;Ewers and Zimmermann,1984).The result of this‘‘bottleneck’’of low conductivity is that under drought condition the wa-
604[Vol.87
A MERICAN J OURNAL OF
B OTANY
ter potential will be lower in the branch than the stem, so growth would stop?rst in the branches and the branch-es will die before the main stem(Zimmermann,1978). Reduced water transport to branches would also reduce the?ow of dissolved nutrients into the branch and there-fore reduce growth.
Several observations suggest that differences in trans-port are the result,not the cause,of apical control.When a young branch of Douglas-?r(Pseudotsuga menzeisii)is released from apical control it bends up and replaces the leader.As the replacement leader grows and produces new xylem its hydraulic conductivity gradually increases, but after15months it is still less than in a leader(Spicer and Gartner,1998).Thus,changes in hydraulic conduc-tivity lag far behind the release from apical control.Phil-lips(1975)pointed out that phloem girdling above a branch does not affect nutrient(or water)transport to the terminal,but does remove apical dominance.This obser-vation also applies to Pinus strobus where phloem girdles above a branch eliminate apical control without affecting terminal growth and presumably water?ow(Wilson and Archer,1981).Girdles below branches also eliminate api-cal control in Pinus strobus.If these girdles had any ef-fect on xylem transport,the hydraulic conductance would be reduced below the branch–stem junction and should have no effect on the relative hydraulic conductance of the stem and branch above the girdle.
Photosynthesis—Older branches appear to be largely autonomous for assimilate and depend on their own pho-tosynthesis for assimilate(Sprugel,Hinckley,and Schaap,1991).Therefore,increased rates of photosyn-thesis after release from apical control could increase branch growth.Leverenz(1981)found that on large branches the?rst-order terminals have higher rates of photosynthesis and transpiration than the higher order lat-erals,even though there was no difference in water po-tential.He did not test the effect of decapitating the ter-minal on photosynthesis in the laterals.The terminal may depresses photosynthesis in the lateral.If this phenome-non occurs,when the terminal is removed the photosyn-thetic rate of the leaves on the lateral would increase. Compensatory increases in photosynthesis can occur after partial defoliation in conifers and angiosperms(Reich et al.,1993;Pinkard et al.,1998).Photosynthesis in re-maining leaves can increase25–75%after partial defo-liation.Although compensatory photosynthesis in a lat-eral after removing apical control has apparently not been demonstrated,it would be consistent with increased growth after removing apical control.
Individual branches in the light grow larger than those in the shade(Stoll and Schmid,1998).These well-lit branches appear to escape some apical control,and they have a higher rate of photosynthesis.Part of the high rate of photosynthesis is due to the higher light intensity and part is due to increased branch growth and leaf area.Pre-sumably the increased photosynthesis permits increased growth,and increased growth may,in turn,increase hor-mone production.
Moderate water or nutrient de?ciencies under apical control would affect growth more than photosynthesis (Cannell and Dewar,1994).Therefore,factors that lead to decreased water or nutrients in branches compared to the terminal could stop branch growth without stopping branch photosynthesis and assimilate production.This as-similate would then be available for export to the ter-minal.
Assimilate allocation—Shoots under apical control all produce leaves,even the short shoots that scarcely elon-gate.Therefore,in contrast to apical dominance where leaves are not produced,all lateral shoots under apical control produce assimilate.The allocation hypothesis is that the parent shoot regulates the export of assimilate from the lateral branch.When the parent axis is intact, assimilate produced by the branch is exported,rather than being used for branch growth.When the parent shoot is removed,the lateral retains assimilate and continues to grow.Ford,Avery,and Ford(1992)assumed for a sim-ulation model that export from the branch began when branch growth requirements had been met.The allocation hypothesis suggests that retention of branch assimilate occurs when the branch sink strength exceeds the ter-minal sink strength.
Experiments with Pinus strobus support the impor-tance of assimilate retention by branches.Pinus strobus has?xed growth.The initial response of the branch to release from apical control is prolonged cambial activity after the branch terminal shoot elongation stops(Wilson and Archer,1981).A stem girdle above a branch removes apical control.Indole-butyric acid(a synthetic auxin)ap-plied to the proximal end of the girdle replaces apical control(Wilson,1986).A girdle2cm below a branch also removes apical control even though the branch is still in direct connection with the terminal(Mu¨nch,1938; Wilson,1981).A girdle50cm below the branch,how-ever,does not remove apical control(Wilson,1981, 1998).My interpretation of these results is that the stem below the branch is a competitive sink for branch-pro-duced assimilate.Auxin moving down the stem and branches by polar transport stimulates cambial activity and the competitive sink strength of the stem.A girdle above the branch stops auxin transport down the stem, reduces stem sink strength,permits the branch to retain assimilate,and removes apical control.Exogenous auxin added above the branch maintains cambial activity and sink strength in the stem below the branch and reimposes apical control.A girdle2cm below the branch makes the stem sink so small that it cannot compete with the branch and apical control is removed.A girdle50cm below the branch makes the stem sink big enough so that the normal levels of auxin moving down the stem create a competitive stem sink and apical control is maintained.
Girdling above a branch removes apical control in all species tested,but the interesting results from Pinus stro-bus are from girdles below branches.Therefore,I at-tempted to repeat the P.strobus experiments on other species.Girdling2cm below branches in the diffuse-porous angiosperms Acer rubrum and Betula lenta did not remove apical control in B.lenta and killed all stem and branch above the girdles in A.rubrum(Wilson, 1998).It is not clear whether apical control is different in these angiosperms or whether the girdles interfere with xylem transport so shoots above the girdle die.
Several observations suggest that branches compete with each other,perhaps by competing for assimilates.
May2000]605
W ILSON—A PICAL CONTROL
Suzuki(1990)thinned lateral buds on parent shoots,and the remaining buds produced longer shoots than in un-thinned controls.Stoll and Schmid(1998)observed that growth of Pinus sylvestris branches in the shade was greater when competing branches were also in the shade than when they were in the sun.They suggested that there was dynamic competition among the branches and that the branches in the sun actually inhibited the growth of those in the shade.This effect would be similar to the effect of apical control,and it could be due to competitive sinks for assimilate.
Gravimorphism—The effects of shoot angle on shoot growth are called gravimorphism(Wareing and Nasr, 1961).The observations are that shoots grow fastest if they are vertical and grow more slowly as their angle to vertical increases.Therefore,branches at an angle to ver-tical would grow more slowly than their more vertical parent shoot just because of gravimorphism.In addition, shoot angle can modify the effect of apical control within and between lateral shoots.Wareing and Nasr(1961) pruned and disbudded cherry,plum,and black currant rootstocks so that there were only two laterals.In controls the uppermost lateral grew the most and was more ver-tical.Bending laterals to horizontal reduced their elon-gation.If only the upper lateral was bent horizontal,the lower lateral elongated the most and grew vertically as a replacement shoot.Thus,a branch bent to horizontal los-es its capacity to control the growth of more proximal branches.Leakey and Longman(1986)found that chang-ing the orientation of rooted cuttings from vertical to hor-izontal reversed the pattern of lateral shoot length from the distal lateral being the longest when vertical to the proximal being the longest when horizontal.
Gravimorphism presumably contributes to the reduced growth of lateral branches that are out of vertical,but why would gravimorphism effects decrease after release from apical control?The branch starts to grow faster be-fore there is any change in angle.Therefore,changes in angle cannot account for initial increases in growth rate, but may be a factor later after the branch bends upward.
APICAL CONTROL OF BRANCH ANGLE Radial growth from cambial activity is necessary,but not suf?cient,for the upward bending of an existing lat-eral branch.Branches of some shrubs respond to remov-ing apical control by increasing diameter growth without upward movement of the branches(Wilson,1998).Al-though plagiotropic branches cannot bend to vertical, both plagiotropic and orthotropic branches of rain-forest angiosperms are more upright in the light than in the shade(King,1998),presumably from increased branch growth in the light.Apical control of conifer branches, measured by the relative elongation of branch and stem terminals,is reduced in the shade,yet branch angle to vertical does not appear to decrease(O’Connell and Kel-ty,1994),probably because radial growth does not in-crease.
Although this section discusses the upward bending of pre-existing woody branches,the geotropic angles of elongating shoots also affect branch angle.The elongat-ing tip of a branch is often more vertical than the older portion of the branch,resulting in an upwardly curved branch,even though there was no upward bending of any woody portions of the branch.
Mechanics—Branches are complex cantilever beams that follow the laws of mechanics(Niklas,1992).A lat-eral shoot at an angle to vertical is always acted on by the downward bending moment from self-mass.The branch cannot bend up unless it can generate enough up-ward bending moment from differential growth stresses in new wood to overcome the downward bending mo-ment.The upward bending moment is a function both of the level of differential growth stress in each wood cell and of the amount of new wood with that differential growth stress.Therefore,from a mechanical point of view,apical control of branch angle can result from reg-ulating the level of differential growth stress in new wood,the amount of new wood with stresses(radial growth),or both.
Some woody plants apparently cannot produce wood with differential growth stresses and therefore their branches cannot bend up.Their only response to release from apical control is increased radial growth(Wilson, 1998).Most woody plants can generate differential growth stresses in wood,usually,but not necessarily,as-sociated with formation of specialized wood(Wilson and Gartner,1996).Upward bending moment apparently can be generated by the bark in some tropical species(Fisher and Mueller,1983).
What is the relative importance of radial growth and the generation of growth stresses?Studies so far suggest that release from apical control always results in an in-crease in branch diameter growth,even in shrubs that cannot generate growth stresses and whose branches can-not bend up(Wilson,1998).Growth stresses in conifer wood are generated primarily by compression wood cells. Compression wood cells are produced in most(not all) branches under apical control(Timell,1986,p.865),but not enough compression wood cells are produced to bend the branch up.After removing apical control of Pinus strobus branches,the level of growth stress was a func-tion of the number of compression wood cells rather than different levels among the cells(Wilson,1986).There-fore,the major effect of removing apical control in co-nifers seems to be to increase radial growth,because compression wood is being formed anyway
Most angiosperms generate upward bending moments by producing tension wood,although bending moments can be generated from apparently normal wood(Wilson and Gartner,1996).In Prunus serotina and Fraxinus americana,branches under apical control formed no ten-sion wood,but those with decapitated terminals formed tension wood and bent up(Wilson and Archer,1983). Therefore,in contrast to conifers,in angiosperms release from apical control may trigger the formation of tension wood along with increased radial growth.
Equilibrium position—The equilibrium position is the most mysterious aspect of branch angle.The equilibrium position is that position where the branch does not pro-duce wood with differential growth stresses.If a branch is bent to either a more(upward),or less(downward), vertical position than the equilibrium position,it will tend
606[Vol.87
A MERICAN J OURNAL OF
B OTANY
to bend back toward the equilibrium position.Therefore, a branch will not bend itself to vertical if the equilibrium position is out of vertical.If the equilibrium position of a shoot is vertical the shoot is called orthotropic,and,if the equilibrium position is out of vertical,the shoot is called plagiotropic.
Some branches are irreversibly plagiotropic and will never bend up to vertical after removal of apical control. The classic example is Araucaria(Timell,1986).The equilibrium position of other plagiotropic branches ap-parently can change after release from apical control. Rooted cuttings of Pseudotsuga menziesii branches,for example,grow plagiotropically at angles to vertical for a year or more before they bend upward to vertical(Star-buck and Roberts,1983).Pinus strobus branches are ini-tially plagiotropic after decapitation of the terminal,but eventually they bend upwards to vertical and become or-thotropic(Wilson,1973).Apparently the equilibrium po-sition of new differentiating wood cells eventually chang-es to vertical in the absence of apical control.Therefore, apical control seems to maintain the equilibrium position of these branches with reversible plagiotropy,but chang-es occur slowly after release from control.
Orthotropic branches may grow at an angle to vertical. If the initial branch angle is out of vertical,there will be a downward bending moment from self-mass.The branch will stay at an angle if it does not grow radially fast enough under apical control to bend upward to vertical. After removing apical control,the radial growth and up-ward bending moment of the branch increase and it can bend upward.
DISCUSSION
Primary mechanisms of apical control should change rapidly after removing apical control.Some factors that regulate branch growth(Fig.2)may be eliminated as primary causes of apical control.For example,the‘‘bot-tleneck’’of decreased hydraulic conductivity at a branch junction can only change gradually as new xylem with higher conductivity is produced.The bottleneck is asso-ciated with branch growth and probably contributes to the relatively slow growth of branches by reducing nu-trients,water potential,and root-produced hormones in the branch,but it is not a primary factor in apical control. Leaf area increase can be eliminated because leaf area does not change until the second year in?xed-growth species,even though cambial activity is prolonged the ?rst year.Direct action of hormones from the terminal on hormone levels in the lateral branch may be rejected,or at least strongly questioned,for Pinus strobus where gir-dles below branches remove apical control even though the connection between the terminal and the lateral are intact.Gravimorphism cannot be involved in initial changes because angle changes are relatively slow.Once the branch starts to bend upward,then reductions in gra-vimorphic effects may permit increases in branch growth.
Compensatory photosynthesis after removing apical control is possible,although there are no data to test whether it https://www.doczj.com/doc/57354638.html,pensatory photosynthesis seems most likely when removing apical control signi?cantly reduces leaf area.Girdling,which does not affect total leaf area,may not result in compensatory photosynthesis,particularly when the girdle is below a branch in Pinus strobus.
Hormone production and assimilate retention by the branch are the most likely candidates for the primary causes of apical control.The two factors are interrelated and are dif?cult to separate.Reduced hormone produc-tion under apical control,or changes in ratios of hor-mones produced,could stop branch growth and stop the branch sink for assimilate.Therefore,removing apical control would prolong hormone production by the branch at promotive levels and prolong the branch sink at com-petitive levels.The question would then become how the controlling parent shoot regulates hormone production by the branch.An alternative mechanism is that reduced as-similates,due to export to the stem,inhibit growth even with adequate hormones.In this case,removing apical control would reduce the strength of the competitive sink in the parent shoot or stem,permit assimilate retention in the branch,and allow continued growth of the branch and production of hormones by the branch.
Branch growth seems to be determined by the relative competitive abilities of the branch and the parent axis.A branch can partially escape apical control if it grows fast enough.Growth of lateral shoots can be increased with high light.The increase could be from more photosyn-thesis and more available assimilate,or it could be from changes in hormone production,or both.Unlike apical dominance,where laterals either grow or do not grow, there is a wide range of levels of apical control under different conditions both between and within individual plants.
Increased branch diameter growth and production of upward bending moments after release from apical con-trol are both important factors in the upward bending of a branch.Increased diameter growth is associated with assimilate retention.The regulation of the formation of growth stresses that cause upward bending moments,for example by reaction wood formation,appears to be a different phenomenon.Timell(1986)thoroughly re-viewed aspects of apical control primarily,but not exclu-sively,of compression wood formation by conifers.He concluded(p.1246)that‘‘the total evidence is too con-tradictory and sometimes downright confusing’’to reach de?nite conclusions.The situation has not changed sig-ni?cantly.The question of how,or even whether,apical control regulates the equilibrium position of a branch, and thus regulates the formation of reaction wood and growth stresses,remains open.
Apical control is so widespread and covers so many aspects of growth that there probably is no single mech-anism for all the phenomena involved.It is possible that different mechanisms act on the central process of assim-ilate allocation.We should be prepared to deal with a complex system.The mechanisms are probably different for the growth processes determining bud size,shoot elongation,and cambial activity.Mechanisms controlling elongation and diameter growth are almost certainly dif-ferent from those controlling growth stress development in wood cells.In addition,there may be differences among various woody species.It is desirable to test hy-potheses and repeat experiments in conifers and angio-sperms,both?xed-and free-growth species.Before try-ing to experiment with basic mechanisms at the cell level
May2000]607
W ILSON—A PICAL CONTROL
we need better descriptions of the phenomena involved in apical control.
LITERATURE CITED
B ORCHERT,R.,AND H.H ONDA.1984.Control of development in the
bifurcating branch system of Tabebuia rosea:a computer simula-tion.Botanical Gazette145:184–195.
B ROWN,C.M.,R.G.M
C A LPINE,AN
D P.P.K ORMANIK.1967.Apical
dominance and form in woody plants:a reappraisal.American Journal of Botany54:153–162.
C ANNELL,M.G.R.,AN
D C.D EWAR.1994.Carbon allocation in trees:
a review of concepts for modeling.Advances in Ecological Re-
search25:59–104.
C HAMPAGNAT,P.1978.Formation of the trunk in woody plants.In P.
B.Tomlinson and M.H.Zimmermann[eds.],Tropical trees as liv-
ing systems,401–422.Cambridge University Press,Cambridge, UK.
C HEN,H.-J.,M.B OLLMARK,AN
D L.
E LIASSON.1996.Evidence that cy-
tokinin controls bud size and branch form in Norway spruce.Phy-siologia Plantarum98:612–618.
C LINE,M.G.1994.The role of hormones in apical dominance:new
approaches to an old problem in plant development.Physiologia Plantarum90:230–237
———.1997.Concepts and terminology of apical dominance.Amer-ican Journal of Botany84:1064–1069.
E MERY,R.J.N.,N.E.L ONGNECKER,AND C.A.A TKINS.1998.Branch
development in Lupinus angustfolius L.II relationship with endog-enous ABA,IAA and cytokinins in axillary and main stem buds.
Journal of Experimental Botany49:555–562.
E WERS,F.W.,AND M.H.Z IMMERMANN.1984.The hydraulic architec-
ture of balsam?r(Abies balsamea).Physiologia Plantarum60: 453–458.
F ISHER,J.B.,AND R.J.M UELLER.1983.Reaction anatomy and reori-
entation in leaning stems of balsa(Ochroma)and papaya(Carica).
Canadian Journal of Botany61:880–887.
F ORD,E.D.,A.A VERY,AND R.F ORD.1992.Simulation of branch
growth in the Pinaceae:interactions of morphology,phenology, foliage productivity,and the requirements for structural support,on the export of carbon.Journal of Theoretical Biology146:15–36.
F ORRESTER,J.W.1968.Principles of systems.Wright-Allen Press,
Cambridge,Massachusetts,USA.
H ALLE′,F.,R.A.A.O LDEMAN,AND P.B.T OMLINSON.1978.Tropical
trees and forests:an architectural analysis.Springer-Verlag,New York,New York,USA.
H OUSE,S.,M.D IETERS,M.J OHNSON,AND R.H AINES.1998.Inhibition
of orthotropic replacement shoots with auxin treatment on decapi-tated hoop pine,Araucaria cunninghami,for seed orchard man-agement.New Forests16:221–230.
K ENNEDY,R.W.,AND J.L.F ARRAR.1965.Induction of tension wood with the anti-auxin2,3,5tri-iodobenzoic acid.Nature208:406–407.
K ING,D.A.1998.The relationship between crown architecture and branch orientation in rain forest trees.Annals of Botany82:1–7. L EAKEY,R.R.G.,AND K.A.L ONGMAN.1986.Physiological,environ-mental and genetic variation in apical dominance as determined by decapitation in Triplochiton scleroxylon.Tree Physiology1:193–207.
L EVERENZ,J.W.1981.Photosynthesis and transpiration in large forest-grown Douglas-?r:interactions with apical control.Canadian Jour-nal of Botany59:2568–2576.
L ITTLE,C.H.A.1970.Apical dominance in long shoots of white pine (Pinus strobus).Canadian Journal of Botany48:239–253.———,AND R.P.P HARIS.1995.Hormonal control of radial and lon-gitudinal growth in the tree stem.In B.L.Gartner[ed.],Plant stems:physiology and functional morphology,281–219.Academic Press,New York,New York,USA.
M OORBY,J.,AND P.F.W AREING.1963.Aging in woody plants.Annals of Botany27:291–308.
M U¨NCH,E.1938.Investigations on the harmony of tree shape.Jahr-bu¨cher fu¨r wissenschaftliche Botanik86:581–673.(Translated by M.H.Zimmermann,1963).N IKLAS,K.J.1992.Plant biomechanics.University of Chicago Press, Chicago,Illinois,USA.
O’C ONNELL,B.M.,AND M.J.K ELTY.1994.Crown architecture of understory and open-grown white pine(Pinus strobus L.)saplings.
Tree Physiology14:89–102.
P HILLIPS,I.D.J.1975.Apical dominance.Annual Review of Plant Physiology26:341–367.
P INKARD,E.L.,C.L.B EADLE,N.J.D AVIDSON,AND M.B ATTAGLIA.
1998.Photosynthesis response of Eucalyptus nitens(Deane and Marden)Maiden to green pruning.Trees12:119–128.
R EICH,P.B.,M.B.W ALTERS,S.C.K RAUSE,D.W.V ANDERKLEIN,K.F.
R AFFA,AND K.F.T ABONE.1993.Growth,nutrition and gas ex-change of Pinus resinosa following arti?cial defoliation.Trees7: 67–77.
R EMPHREY,W.R.,AND C.G.D AVIDSON.1992.Branch architecture and its relation to shoot tip abortion in mature Fraxinus pennsylvani-cum.Canadian Journal of Botany70:1147–1153.
R ICHARDS,J.H.,AND P.R.L ARSON.1981.Morphology and develop-ment of Populus detoides branches in different environments.Bo-tanical Gazette142:382–393.
S CHMITZ,G.,AND K.T HERES.1999.Genetic control of branching in Arabidopsis and tomato.Current Opinion in Plant Biology2:51–
55.
S PICER,R.,AND B.L.G ARTNER.1998.How does a gymnosperm branch (Pseudotsuga menseisii)assume the hydraulic status of a main stem when it takes over as a leader?Plant,Cell and Environment21: 1063–1070.
S PRUGEL,D.G.,T.M.H INCKLEY,AND W.S CHAAP.1991.The theory and practice of branch autonomy.Annual Review of Ecology and Systematics22:309–322.
S TARBUCK,C.J.,AND A.N.R https://www.doczj.com/doc/57354638.html,pression wood in rooted cuttings of Douglas-?r.Physiologia Plantarum57:371–374. S TOLL,P.,AND B.S CHMID.1998.Plant foraging and dynamic compe-tition between branches of Pinus sylvestris in contrasting light en-vironments.Journal of Ecology86:934–945.
S UNDBERG,B.,AND C.U GGLA.1998.Origin and dynamics of indole-acetic acid under polar transport in Pinus sylvestris.Physiologia Plantarum104:22–29.
S UZUKI,T.1990.Apical control of lateral bud development and shoot growth in mulberry(Morus alba):effects of position of lateral and accessory buds and leaves.Physiologia Plantarum80:350–356. T HIMANN,K.V.1977.Hormone action in the whole life of plants.
University of Massachusetts Press,Amherst,Massachusetts,USA. T IMELL,https://www.doczj.com/doc/57354638.html,pression wood in gymnosperms.Springer-Verlag,New York,New York,USA.
W AREING,P.F.,AND T.A.A.N ASR.1961.Gravimorphism in trees I.
Effects of gravity on growth and apical dominance in fruit trees.
Annals of Botany25:321–340.
W ILSON,B.F.1973.White pine shoots:roles of gravity and epinasty in movements and compression wood location.American Journal of Botany60:597–601.
———.1981.Apical control of diameter growth in white pine branch-es.Forest Science27:95–101.
———.1986.Apical control of compression wood action in white pine branches.Wood Science and Technology20:111–117.———https://www.doczj.com/doc/57354638.html,pensatory growth in shoot populations of young white pine trees.Trees6:204–209.
———.1998.Branches versus stems in woody plants:control of branch diameter growth and angle.Canadian Journal of Botany 76:1852–1856.
———,AND R.R.A RCHER.1981.Apical control of branch movements in white pine:biological aspects.Plant Physiology68:1285–1288.———,AND———.1983.Apical control of branch movements and tension wood in black cherry and white ash trees.Canadian Jour-nal of Forest Research13:594–600.
———,AND B.L.G ARTNER.1996.Lean in red alder(Alnus rubra): growth stress,tension wood and righting response.Canadian Jour-nal of Forest Research26:1951–1956.
———,AND M.J.K ELTY.1994.Shoot growth from the bud bank in black oak.Canadian Journal of Forest Research24:149–154.
Z IMMERMANN,M.H.1978.Hydraulic architecture of some diffuse-po-rous species.Canadian Journal of Botany56:2286–2295.
慈善组织嫣然天使基金 2019-01-01 嫣然天使基金(Smile angel Foundation )是由李亚鹏、王菲倡导发起,在中国红十字基金会的支持和下设立的专项公益基金,2006年11月21日正式启动, 。 2014年2月红十字总会委托会计师事务所开始审计李亚鹏的嫣然基金账目。 2014年8月12日,民政部回应,通过对中国红十字会总会及会计师事务所提供的材料进行研究核查,没有发现举报人所举报的问题。 基本介绍: 中国红十字基金会将与发起人共同倡导、动员社会资源,以弘扬人道、博爱、奉献的红十字精神为宗旨,积极为嫣然天使基金筹集医疗救助资金,让更多唇腭裂患儿得到治疗,拥有一张纯真的笑脸。 据有关部门统计,中国有240万唇腭裂患者。唇腭裂是最常见的先天畸形之一。根据中国出生缺陷检测中心对全国31个省、市、自治区466所医院每年40万-70万新生儿的检测结果显示,1988年-1992年非综合性唇腭裂发生率为1.625‰,其中唇裂发生率为1.4‰、腭裂发生率为0.225‰。在此背景下,嫣然天使基金的成立具有十分重大的意义。 嫣然天使基金的资助对象为家庭贫困身患唇腭裂的.患者,患儿法定监护人可作为申请人向嫣然天使基金申请资助, 《》()。具体申请方法为:登陆中国红十字基金会、中国重症儿童救助网,下载嫣然天使基金资助申请表,在完全理解《嫣然天使基金申请须知》的前提下,经县级以上红十字会审核盖章后,向中国红十字基金会提出求助申请,并提交资料。嫣然天使基金资助管理办公室对患儿的申请资料进行初审后,会同定点医院对患儿进行全面的术前检查、排除手术禁忌症、部分患儿会诊,形成基本的医治方案,连同患儿法定监护人签署的手术知情同意书一并提交嫣然天使基金管理委员会审批。嫣然天使基金管理委员会根据资助原则对患儿进行综合评审,确定资助对象和资助金额。获资助对象名单将在中国红十字基金会进行公示。患儿凭中国红十字基金会嫣然天使基金资助通知书到指定医院入院治疗。 儿童是社会的未来,儿童是祖国的希望,关心唇腭裂儿童是全社会的责任。希望社会各界广发博爱之心,共同关注嫣然天使基金! 成立背景:
融资担保公司工作总结 导读:范文融资担保公司工作总结 【范文一:诚信融资担保公司工作总结】 20XX年公司在县委、县府的关心支持下,在县国资委直接领导和指导下,公司坚持诚实守信,控制风险的经营原则,积极、稳妥地开展工作,充分发挥融资担保杠杆作用,有力地解决和缓解了我县中小企业、个体经营者的生产、经营资金的困难,大力支持了我县中小企业的发展,取得了较好的社会效益。 一:担保业务及资产经营情况 1、担保业务工作 20XX年由于国家宏观经济调控,实行适度稳健的货币政策,压缩银行贷款,我公司为了加强贷款风险管理,降低融资担保风险,采取了压缩融资担保额度的经营策略。20XX年公司累计担保132笔,担保金额16916万元,年末在保责任余额18980万元,其中:在保企业48户,在保余额17870万元、在保个人30户,在保余额1110万元。
3、担保代偿及追偿工作 20XX年公司累计担保代偿27笔,金额1544万元,其中:为华祥集团提供的银丰国际中心项目工程款履约担保代偿及贷款担保代偿960余万元。 通过法院起诉15笔,金额1200余万元,已通过法院执行、调解追偿收回189余万元。其中:华祥集团提供的银丰国际中心项目工程款履约担保代偿及贷款担保代偿960余万元正在执行过程之中。 3、资产经营情况: 20XX年末公司资产总额6702万元,较投入的国有资本总额5000万元增加1702万元,增值34%。负债总额1355万元,所有者权益5347万元。20XX年,公司实现保费收入173万元,净利润1、2万元。 二、主要工作开展情况: 1、努力拓展担保业务,最大程度缓解企业融资难问题 作为国有控股的政策性担保公司,我们始终以促进县域经济发展,提高中小企业信用,拓宽中小企业融资渠道,推动中小企业改革与发
嫣然基金 ——蒋婕091504110 创始人:王菲李亚鹏夫妇 2006年11月21日,嫣然天使基金正式启动。该基金是由 李亚鹏、王菲夫妇倡导发起,在中国红十字基金会的支持和管理 下设立的专项公募基金。 背景介绍:目前,中国有240万名唇腭裂孩子,不少家庭贫困的 唇腭裂儿童没钱治疗,只能以一个残缺的形象面对社会和世人。 根据我国出生缺陷检测中心对全国31个省、市、自治区466所 医院每年40万-70万新生儿的检测结果显示,1988年-1992 年非综合性唇腭裂发生率为1.625‰,其中唇(腭)裂发生率为 1.4‰、腭裂发生率为0.225‰。针对这种情况,李亚鹏表示:“这 是我最想去帮助的一个群体。” 为了帮助这些孩子,李亚鹏、王菲夫妇慷慨捐款100万,“嫣 然天使基金”应运而生——以个人名义发起的爱心基金,在中国 红十字基金会还是第一个。 嫣然大事记: 2006年9月李亚鹏王菲夫妇向中国红十字基金会提交了申请,并捐献100万元作为启动资金。 2007年12月8日第二届爱心慈善晚宴在上海外滩的茂悦酒店举行,共筹款2029万元。 2008年“5 ·12汶川地震”后,拨款500万,特别用于救助四川
汶川地震中受伤的儿童,专门成立专项救助医疗队旨在为灾后受损儿童进行整形修复、功能修复、瘢痕修复和器官再造等治疗。 2009年9月29日至10月7日,李亚鹏亲自带领嫣然天使基金医疗队远赴西藏阿里救助唇腭裂儿童,此次救助被称为是建国以来“历时最长、海拔最高、医生数量最多”的一次慈善救助行动,共为34名唇腭裂患者成功手术。 2009年12月18日第三届爱心慈善晚宴在北京柏悦酒店举行,共筹得善款现金2956万元,北大药业物资5000万元,共计7956万元。组织特点: 资助对象 嫣然天使基金的资助对象为家庭贫困身患唇腭裂的患者,患者法定监护人可作为申请人向嫣然天使基金申请资助。 资助原则 1.量入为出:根据接收捐款情况确定资助名额; 2.全额资助:承担全部医疗费用; 3.功能恢复:只负担唇腭裂患者唇腭裂的功能恢复缝合手术。 申请程序 1.申请人需通过中国红十字基金会官方网站下载嫣然天使基金求助申请表,在完全理解《嫣然天使基金申请须知》的前提下,填写表格(包括以下内容): (1)患者身份证明文件复印件; (2)如属患儿,还需提供患儿法定监护人的关系证明文件;
《服务指南》 事项名称:融资担保公司设立审批(跨省、自治区、直辖市设立分支机构)(行政许可) 适用范围:申请在防城港市区内设立分公司的融资担保公司 事项审查类型:前审后批 三、审批依据: 1. 国务院《融资担保公司监督管理条例》(2017年6月21日国务院第177次常务会议通过,由中华人民共和国国务院于2017年8月2日发布,自2017年10月1日起施行)第六条设立融资担保公司,应当经监督管理部门批准。 2. 《国务院对确需保留的行政审批项目设定行政许可的决定》(2004年6月29日国务院令第412号发布,2016年8月25修订,自2004年7月1日起施行)附件《国务院对确需保留的行政审批项目设定行政许可的决定》(2004年6月29日国务院令第412号发布,2016年8月25修订,自2004年7月1日起施行)附件第十二项“跨省区或规模较大的中小企业信用担保机构设立与变更审批”,《中华人民共和国国务院令》第
548号(2009年1月29日发布)一、将第12项的项目名称,由“跨省区或规模较大的中小企业信用担保机构设立与变更审批”修改为“融资性担保机构的设立与变更审批”;将实施机关,由国家发展改革委改为省、自治区、直辖市人民政府确定的部门。 四、受理机构:防城港市行政审批服务中心金融办窗口 五、决定机构:自治区地方金融监督管理局融资担保公司监管处 六、数量限制:无数量限制 七、申请条件: 根据《广西壮族自治区融资性担保公司管理暂行办法》(桂政发〔2010〕49号)第十四、十五条和《广西融资性担保公司设立申报指引(2015年版)》第二大点的相关规定,设立融资性担保分公司应当具备下列条件:设立分公司 (一)区内的融资性担保公司设立分公司 1.经营融资担保业务一年以上,注册资本不少于人民币1亿元,放大倍数5倍以上。 2.稳健、合规经营,最近2年盈利。
信用担保机构助力中小企业融资对策研究 近年来,中小企业对我国的GDP和就业作用巨大。伴随着其快速发展,融资需求越来越大,对信用担保机构的依赖程度也就越强。信用担保机构打破了传统的双方交易模式,弥补了传统融资行业的不足,扩展了融资交易范围,增加了中小企业的受保范围,打破了以往中小企业融资的局限性,使因自身条件导致从银行贷 款或从银行无法得到满足自身资金需求的中小企业获得贷款,对促进中小企业融资具有重要的现实意义。信用担保机构是维系中小企业与银行关系牢固的桥梁,是促进双方共同发展的关键纽带,很好地助力了中小企业融资。 1993年中国首家信用担保机构成立,标志着中国担保业开始快速发展,随后 信用担保机构大量出现。但随着发展速度的加快,信用担保机构出现了许多问题,例如:信用担保机构的担保合同设计不标准、资本金少、专业人才少、创新不足等。这些问题完全涵盖整个信用担保体系,影响了中小企业融资进程。同时随着越来越多的中小企业通过信用担保机构进行融资,融资时所面临的困难也逐渐增多。 为此,针对中小企业融资难、难融资现象,开始逐步完善信用担保体系,不断寻求更多的对策,如提升信用等级、降低信息不对称、增加贷款额度等来助力中小企业融资。在当前大数据、互联网及普惠金融的背景下,信用担保机构如何更好地助力中小企业融资已成为当前急需探讨的问题。本文从信用担保机构视角入手,对中小企业融资对策进行研究。第一章,运用文献综述法对中小企业融资及信用担保机构的国内外文献进行了简要介绍,系统地了解了国内外学者在信用担保机构助力中小企业融资方面所取得的优秀研究成果。 第二章,对信用担保机构的核心概念及基础理论进行了阐述。第三章,简要介绍了我国信用担保机构助力中小企业融资的现状及所遇到的问题。第四章,深入探讨信用担保机构对中小企业融资的作用机制,如提升中小企业融资增信能力及信用等级,降低中小企业融资的信息不对称,提高抗风险能力,完善监管服务体系。第五章,运用案例分析法,选取中国投融资担保股份有限公司作为国内信用担保 机构的典型代表,剖析其当前助力中小企业融资时采用的业务模式以及所取得的业绩。 第六章,结合第三章信用担保机构助力中小企业融资时遇到的问题和第五章
证券代码:688007 证券简称:光峰科技公告编号:2020-015 深圳光峰科技股份有限公司 关于2020年度公司及子公司申请综合授信额度 并提供担保的公告 重要内容提示: ●2020年度公司及子公司拟向银行等金融机构申请不超过36亿元的综合 授信额度,并为综合授信额度内的子公司融资提供不超过16.2亿元的 担保额度。 ●被担保人:控股子公司中影光峰激光影院技术(北京)有限公司和峰米 (北京)科技有限公司,全资子公司光峰光电香港有限公司 ●截至本公告披露日,公司对子公司已实际发生的担保余额为29,548万 元。 ●被担保人未提供反担保,公司无逾期对外担保情形 ●本事项尚需提交公司2019年年度股东大会审议 一、2020年度申请综合授信额度并提供担保情况概述 (一)情况概述
为满足经营和发展需求,公司及子公司2020年度拟向银行等金融机构申请不超过人民币36亿元的综合授信额度,主要用于办理流动资金贷款、固定资产贷款、合同融资、并购贷款、开立银行承兑汇票、信用证、保函、汇票贴现、银行保理业务、贸易融资、供应链融资等业务品种,具体业务品种、授信额度和期限以各家金融机构最终核定为准。以上授信额度不等于公司的实际融资金额,实际融资金额在总授信额度内,以公司及子公司与金融机构实际发生的融资金额为准。 为满足子公司经营和发展需求,提高公司决策效率,公司拟为控股子公司中影光峰激光影院技术(北京)有限公司(以下简称“中影光峰”)、控股子公司峰米(北京)科技有限公司(以下简称“峰米科技”)、全资子公司光峰光电香港有限公司(以下简称“光峰香港”)就上述综合授信额度内的融资分别提供不超过人民币10亿元、5.2亿元、1亿元的担保额度,合计不超过人民币16.2亿元的担保额度,担保方式包括保证、抵押、质押等,具体担保期限根据届时签订的担保合同为准。 公司董事会提请股东大会授权公司经营管理层根据公司实际经营情况的需要,在上述综合授信额度及担保额度范围内,全权办理公司向金融机构申请授信及提供担保相关的具体事项。 (二)审批程序 公司于2020年4月28日召开的第一届董事会第二十一次会议以7票赞成、0票弃权、0票反对的表决结果审议通过了《关于2020年度公司及子公司申请综合授信额度并提供担保的议案》。独立董事对本次授信及担保事项发表了明确同意的独立意见。
"嫣然天使基金”求助申请表 患者照片 (唇裂患者提供正面照片,腭裂患者提供口腔内部照片) 编号:No.: 患者:_________ 性别:—出生日期:年月日 通信地址:________ 省______ 市(县)______________________ 联系人:_____________ 、手机: ______________________ 患者体重:________________ 邮编:______________________ 电子:________________________________________________ 申报日期:
申报须知 1."嫣然天使基金”资助申请表由中国红十字基金会“嫣然天使基金”办公室印制 并负责解释; 2.“嫣然天使基金”资助对象为贫困家庭的唇腭裂患者; 3.患者的所有申报资料由患者或患者的法定监护人负责填报,并保证所有资料的 真实性和完整性; 4.本申报表的递交并不表明肯定获得资助; 5.“嫣然天使基金”办公室负责所有申报资料的审核和建档工作; 6.得到嫣然天使基金资助的唇腭裂患者,嫣然天使基金承担患者的全部医疗费 用。医院提供一名患儿家长陪床的床位。 7.对申报资料中出现的虚假、伪造或隐瞒等行为,一经发现,评审办公室将不予 资助。 8.获得资助的患者或患儿监护人均有责任和义务为评审办公室提供必要的文字、 照片、影像等资料,配合评审办公室的宣传和采访活动,并同意使用其照片、影像等资料。 我确认已经阅读和知悉了以上全部条款,并同意所有申报规定。 患者或患儿监护人签字: 年月日 申请登记表
患者医疗情况简述 患者在治疗过程中的介绍:(请参考后面的附注说明,尽可能详细一些) 1.现在是否有先天性心脏病、传染病等? 2.唇腭裂是否接受过手术治疗?在哪家医院进行治疗?效果如何? 3.目前有无上呼吸道感染、发热及胸腺肥大等情况? 4.目前体重是多少?
ⅩⅩ市小微企业融资担保有限公司 反担保管理办法 第一章总则 第一条为加强对公司担保业务设定的反担保的有效管理,规范公司反担保行为,提高第二还款来源防范风险的能力,根据《中华人民共和国民法通则》、《中华人们共和担保法》等有关法律、法规以及公司有关制度规定,制定本办法。 第二条本办法所称反担保,系指本公司为债务人向债权人提供担保时,为保证本公司承担责任后债权不致悬空,应本公司的要求债务人或第三人向本公司提供的担保。 本办法所称反担保管理,系指本公司通过对反担保的方式和内容、反担保资产的评估、反担保物的登记、保险和公证、反担保资产的管理和处置等内容的全面、系统的管理,规范反担保行为,切实发挥担保防范反担保风险的作用,提高公司控制风险的能力。 第三条本公司接受债务人委托向债权人提供信用担保服务时,均应要求债务人或第三人向本公司提供反担保,公司特许批准的除外。 第四条根据国家法律的规定和担保行业所面临的现实环境,本公司担保业务的反担保措施为保证、抵押、质押以及其他商业化反担保方式。 公司根据担保行业的特点和担保企业的特点,针对每笔担保业务的实际情况,确定包括多种担保方式在内的反担保组合方案,以强化反担保对债务人的威慑作用,保证本公司债权的现实。 公司设定反担保,一般以被担保企业有形资产的抵(质)方式为主,以无形资产、权利(益)质押及其他担保方式为辅。对于反担保
较弱的中小企业,应要求其法定代表人个人对公司债务承担无限连带责任。 第五条公司设定反担保应遵循五项原则: (一)合法、合规原则,即反担保物为国家法律允许设定担保的资产; (二)流通可变现原则,即反担保物为市场所接受,具有广泛的流通性,可通过市场变现; (三)市场定价原则,即通过市场确定反担保物的真实价值; (四)执行可操作原则,即按照相关法律、法规的形式设定反担保物,以保证反担保物能够顺利处置; (五)债务人利益可触动原则,即反担保物的设定,必须能够触动债务人的切身利益,以迫使债务人守信履约。 第六条本办法适用于公司各类担保业务,适用于公司各部门。 第二章反担保的方式和内容 第一节反担保保证 第七条反担保保证是指本公司与反担保保证人约定,当被保证人不能履行债务时,反担保保证人按照约定履行债务的行为。 第八条反担保保证人可分为两类,一是法人或者其他经济组织,二是自然人。 第九条法人或者其他经济组织承担作为反担保保证人应具备下列条件: (一)持有经过年检的法人营业执照、组织机构代码、税务登记证; (二)通过注册有固定的经营场所;
西安高新技术开发区融资情况调研报告 科技部研究中心创业投资研究所郭谦和郭戎 【摘要】经过12年的发展,西安高新区的各项经济指标在全国53个国家级高新区的名列前茅,并首批加入亚太经合组织科技工业园区网络。本文对西安高新区在扶持科技型中小企业融资方面独特的方法、灵活和高效的机制以及西安高新区的经验中体现出的一些值得借鉴的、带有共性的规律进行了分析和总结。 一、西安高新区的特点和基本情况 1.自然和区位情况 西安高新区创建于1991年6月,位于西安市科研、文教单位相对集中的南郊。总规划面积达97.7平方公里,已完成15平方公里的开发建设。区内建设六个专业园区、四个大学园区和长安科技产业基地,形成“十园一基地”的产业发展格局。 2.独特的产业发展模式 西安高新区地处中国腹地,物流是其短边,需要找准位置,充分挖掘自身优势,创造出不同于北京、苏州高新区的发展模式,才可能有所作为。北京中关村高新区首先是IT贸易集散地,然后形成了联想、方正等一批民营企业,走的是贸工技发展道路。苏州高新区利用毗邻上海的地理优势,大规模吸引国外制造业,形成制造业基地和产业群。政府通过对跨国公司的引进获取原始积累,并在此基
础上希望培育出水平相对较高,附加价值较高的创新能力。相比而言,西安既不具有市场优势,也不具有地理优势,模仿北京、苏州高新区的成功模式显然行不通。因此西安高新区充分利用当地科研院所、大学里技术人员的科技资源优势,通过科技人员创办企业,实现科技成果转化,大力发展具有自主知识产权的高科技产业,形成了电子信息、生物医药、光机电一体化三大支柱产业,涌现了一批知名企业,如大唐电信、庆安冷机、兄弟标准、开米股份、协同软件、海天天线、康鸿电子等。 3.强大的区内企业 西安高新区目前有科技企业4991家,股份制企业128家(其中上市公司18家);其中高新技术企业虽然只占企业总数的15.1%,但是技工贸总收入、工业总产值却占全区的75.56%和76.4%,利润、税收占80.41%和76.4%,外资企业582家,占到约10%,世界500强投资的企业20多家。2002年,高新区完成技工贸总收入481.17亿元,工业总产值318.43亿元,GDP120.53亿元,已连续五年拉动西安经济增长超过4个百分点,成为陕西、西安最大的经济增长点和对外开放的窗口。 就企业规模而言,2002年收入过20亿元的企业5家,过10亿元的企业10家,过亿元的企业70家(居全国高新区第4位),过1000万元的企业有295家。大中型企业技工贸总收入和总产值分别占到全区的80.05%和77.54%。2002年销售收入过亿元的产品25个,过5000万元的产品60多个,且大部分具有自主知识产权。高新技术企业销售收入占西安高新区总收入的82%,拥有自主知识的产品销售收入占高新技术企业销售收入的81%。 二、西安高新区独特的融资体系 西安高新区1991年起步时只有10万元的建设资金,而且以上材料表明,西安的科技企业不是由外资企业创办的,所以实力较小,风险很大。这就意味着政府必须在较高的风险不能为市场所化解的情况下,介入扶持科技企业的成长过程。西安高新区之所以最终取得了很大的成功,关键得益于培育、建立了一个高效的、多元化、网络化投融资体系,坚持“政府引导、市场运作”,健全资本市场机制方面所做的创新之举。 1.政府为科技企业成长的各阶段中量体定做的融资方案 企业的发展史就是融资史。对于一个产品,企业总是经历初创期、快速成长期、成长期、稳定期和衰退期,每个阶段,企业的资产状况不同,就需要不同的融资渠道为之服务。但是我国只有单一的银行贷款渠道,显然不能满足处于初创期和快速成长期的企业的资金饥渴。针对这样的状况,西安高新区为企业量身订做不同的融资渠道: ◆在企业初创期,即科研开发阶段,高新区创办了种子资金,由西安创新投
ABC有限公司U0YY年度报告
目录 第一部分公司概况.。。。。。。 一、公司简介。 二、经营计划。 三、组织架构、分支机构设置及人员情况。 四、合作的金融机构 第二部分公司治理和内部控制 一、公司股东、基本情况及报告期内变动情况。 二、U0YY年度内召开始股东会及重要决议。 三、董事会的构成及其工作情况。 四、监事会的构成及其工作情况。 五、高级管理层的构成及其基本情况。 六、内部控制情况 第三部分风险管理 一、风险管理概况 二、信用风险管理 三、流动性风险管理 四、市场风险管理 五、操作风险管理 六、其他风险管理 第四部分担保业务总体情况 一、担保业务总体情况表: 二、集中度情况表: 三、其他情况说明: 四、监管部分检查及整改情况 第六部分资本金构成及资金运用 第七部分财务报告
第一部分公司概况一、公司简介 (一)、公司基本信息
(二)、U0YY年度公司基本财务和业务信息 单位:万元 二、经营计划 U0YU年在金融环境变好的情况下,公司预计将增加担保业务U000万元。公司准备增加乡镇网点Y-U个。公司预计增资U000万元,充实公司资本。 三、组织架构、分支机构设置及人员情况 (一)、组织结构图
(二)、分支机构设置(无) (三)、人员情况 四、合作的金融机构 江苏宿迁HHHHH农村商业银行(以下简称HHHHH银行)是宿迁市区唯一一家独立的地方性法人金融机构,也是市区服务网点较多、分布范围较广、服务功能完备的金融机构。 江苏GGGGGG村镇银行位于江苏TYYY区,是由UUU银行、其他非金融机构企业法人和自然人共同出资设立的地方性银行业金融机构,注册资金K000万元,实行自主经营、自担风险、自负盈亏,以服务三农和广大地方中小企业及区域经济为经营方向。
“嫣然天使基金”资助申请表 并负责解释; 2. “嫣然天使基金”资助对象为 0-14周岁贫困的唇腭裂儿童; 3?患者的所有申报资料由患者的法定监护人负责填报,并保证所有资料的真实性和 完整性; 4. 本申报表的递交并不表明肯定获得资助; 5. 评审办公室负责所有申报资料的审核和建档工作; 6?得到嫣然天使基金资助的唇腭裂儿童,嫣然天使基金承担患者的全部医疗费用, 不承担患 者的路费、餐费。医院提供一名患儿家长陪床的床位。 7. 对申报资料中出现的虚假、伪造或隐瞒等行为,一经发现,评审办公室将不予资 助。 8. 获得资助的患者监护人均有责任和义务为评审办公室提供必要的文字、照片、影 像等资 患者5寸彩照(唇裂照正面照片1张,号腭裂照口腔内照片o. 1张)粘贴处 申
料,配合评审办公室的宣传和采访活动,并同意使用其照片、影像等资料。
我确认已经阅读和知悉了以上全部条款,并同意所有申报规定 患者监护人签字: 年月日 申请登记表
患者医疗情况简述 患者在治疗过程中的介绍(回答如下问题):(请参考后面的附注说明,尽可能写详细- 些) 1. 现在是否有先天性心脏病、传染病等? 2. 孩子唇腭裂是否接受过手术治疗?在哪家医院进行治疗?效果如何? 3. 孩子目前有无上呼吸道感染、发热及胸腺肥大等情况?(当地医院出具检查报告) 4. 孩子目前体重、血常规(白细胞、血色素)怎样?(当地医院出具检查报告) 患者申请嫣然天使基金免费手术资助的原因是什么? 附件一身份证明
患者的户口复印件或出生证复印件(用A4纸复印后装入该申请材料中): 患者父母(或监护人)身份证和户口复印件(用A4纸复印后装入该申请材料中):
嫣然天使基金申请须知 一、资助对象 嫣然天使基金的资助对象为家庭贫困身患唇腭裂的患者,患者法定监护人可作为申请人向嫣然天使基金申请资助。 二、资助原则 1.量入为出:根据接收捐款情况确定资助名额; 2.全额资助:承担全部医疗费用; 3.功能恢复:只负担唇腭裂患者唇腭裂的功能恢复缝合手术。 三、申请程序 1.申请人需通过中国红十字基金会官方网站下载嫣然天使基金求助申请表,在完全理解《嫣然天使基金申请须知》的前提下,填写表格(包括以下内容): (1)患者身份证明文件复印件; (2)如属患儿,还需提供患儿法定监护人的关系证明文件; (3)患者或患儿法定监护人填写的嫣然天使基金资助申请表; (4)患者在县级以上医疗机构的初步体检报告; (5)持非农业户口的2岁以下的贫困家庭的唇腭裂患儿,需在城镇街道办事处以上行政机构出具的家庭经济状况证明。 2.申请资料报送中国红十字基金会嫣然天使基金项目管理办公室。 四、体检 患者或患者法定监护人在完全理解嫣然天使基金资助告知书的内容后,需到当地县级以上医疗机构进行与唇腭裂手术相关的初步检查,并出具体检报告。以下为唇腭裂手术禁忌症,体检报告应涵盖以下内容: 1.唇裂的手术禁忌症: (1)患儿的体重少于5㎏; (2)血红蛋白低于10g/100ml; (3)白细胞计数高于104/mm3或凝血功能异常; (4)患儿的年龄小于10周; (5)患者有急性感染、感冒、上呼吸道感染; (6)患者有消化道疾病; (7)面部、口周及耳鼻咽喉部有炎症疾患; (8)扁桃体过大可能影响手术后呼吸者; (9)患者不能耐受全麻手术。
2.腭裂的手术禁忌症: (1)患儿的体重少于5㎏; (2)血红蛋白低于10g/100ml; (3)白细胞计数高于104/mm3或凝血功能异常; (4)患儿的年龄小于10周; (5)患儿有急性感染、感冒、上呼吸道感染; (6)患者有消化道疾病; (7)面部、口周及耳鼻咽喉部有炎症疾患; (8)扁桃体过大可能影响手术后呼吸者; (9)患者胸腺肥大; (10)患者不能耐受全麻手术。 五、审批程序 1.初审 嫣然天使基金资助管理办公室对患者的申请资料进行初审后,会同定点医院对患者进行全面的术前检查、排除手术禁忌症、部分患者会诊,形成基本的医治方案,连同患者或患者法定监护人签署的手术知情同意书一并提交嫣然天使基金管理委员会审批。 2.审批 嫣然天使基金管理委员会根据资助原则对患者进行综合评审,确定资助对象和资助金额。 3.公示 获资助对象名单将在中国红十字基金会官方网站进行公示。 六、入院治疗 患者接到中国红十字基金会嫣然天使基金办公室电话通知后,到指定医院入院治疗。 七、特别约定 因嫣然天使基金为患者提供的是慈善手术,医患双方特作此约定。定点医院为患者提供免费治疗的时间段截止至患者的病情相对稳定或患者符合出院指征。且该时间段由定点医院根据患者的病情提出建议,嫣然天使基金管理委员会作出最终决定。 八、最终解释权 以上所有条款最终解释权属嫣然天使基金管理委员会。
深圳担保公司名单深圳市中小企业信用融资担保集团有限公司 深圳市华融融资担保有限公司 许可证编码:粤88002 深圳中科智融资担保有限公司 许可证编码:8800258) 深圳市中兰德融资担保集团股份有限公司 许可证编码:8800259) 深圳市融捷融资担保有限公司 许可证编码:粤8800260) 深圳市金瑞格融资担保有限公司 许可证编码:粤8800261) 深圳市中科创融资担保有限公司 许可证编码:8800262) 深圳安信巨融融资担保有限公司 许可证编码:粤8800263) 深圳市利钊融资担保有限公司 许可证编码:粤8800264) 深圳市中锦融资担保有限公司 许可证编码:粤8800265) 深圳市世银联融资担保集团有限公司 许可证编码:8800266) 深圳中联银融资担保有限公司 许可证编码:粤8800267) 深圳市信融通融资担保有限公司 许可证编码8800268) 深圳市二十一世纪融资担保有限公司 许可证编码:粤8800269) 深圳市星日融资担保有限公司 许可证编码:粤8800270) 深圳市中金创展融资担保股份有限公司 许可证编码:粤8800271) 深圳市羽邦融资担保股份有限公司 许可证编码:粤8800272) 深圳市润通融资担保有限公司 许可证编码:粤8800273) 深圳市中合融资担保有限公司 许可证编码:粤8800274) 深圳市达通融资担保有限公司 许可证编码:粤8800275) 深圳市融浩融资担保有限公司 许可证编码:粤8800276) 深圳市中融信融资担保有限公司 许可证编码:粤8800277)
许可证编码:粤8800278) 深圳市智通融资担保有限责任公司 许可证编码:粤8800279) 深圳经纬盈富融资担保有限公司 许可证编码:粤8800280) 深圳市力合智通融资担保股份有限公司许可证编码:粤8800281) 深圳深业融资担保有限公司 许可证编码:粤8800282) 深圳市盛世海基融资担保有限公司 许可证编码:粤8800283) 深圳市光电行业融资担保有限公司 许可证编码:粤8800284) 深圳市富海荣基融资担保有限公司 许可证编码:粤8800285) 深圳市华融达融资担保有限公司 许可证编码:粤8800286) 深圳市中信嘉华融资担保有限公司 许可证编码:粤8800287) 深圳市诺信融资担保有限公司 许可证编码:粤8800288) 深圳市银盛融资担保有限公司 许可证编码:粤8800289) 深圳市不动产融资担保股份有限公司许可证编码:粤8800290) 深圳市诺亚信融资担保有限公司 许可证编码:粤8800291) 深圳市深航融资担保有限公司 许可证编码:粤8800292) 深圳市通银融资担保有限公司 许可证编码:粤8800293) 深圳市中诚致信融资担保有限公司 许可证编码:粤8800294) 深圳长城融资担保控股有限公司 许可证编码:粤8800295) 深圳市立信融资担保有限公司 许可证编码:粤8800296) 深圳市创锋融资担保有限责任公司 许可证编码:粤8800297) 深圳市盛泽融资担保有限责任公司 许可证编码:粤8800298) 深圳市兴业融资担保有限公司 许可证编码:粤8800299)
100301134 代玉婷100301136 周霞100301138 赵姝玉
?嫣然天使基金是由李亚鹏、王菲夫妇于2006年11月21倡导发起,在中国红十字基金会的支持和管理下设立的专项公益基金,旨在救助家庭贫困的唇腭裂儿童。 ?目前,中国有240万名唇腭裂孩子,不少家庭贫困的唇腭裂儿童没钱治疗,只能以一个残缺的形象面对社会和世人。?为了帮助这些孩子,李亚鹏、王菲夫妇慷慨捐款100万,“嫣然天使基金”应运而生——以个人名义发起的爱 心基金,在中国红十字基金会还是第一个。 ?中国红十字基金会将与发起人共同动员社会资源,为嫣然天使基金筹集医疗救助资金,让唇腭裂患儿拥有一张纯真的笑脸。
?资助对象:嫣然天使基金的资助对象为家庭贫困身患唇腭裂的患者,患者法定监护人可作为申请人向嫣然天使基金申请资助。 ?资助原则:1.量入为出:根据接收捐款情况确定资助名额; 2.全额资助:承担全部医疗费用; 3.功能恢复:只负担唇 腭裂患者唇腭裂的功能恢复缝合手术。 ?申请程序:1.申请人需通过中国红十字基金会官方网站下载嫣然天使基金求助申请表,在完全理解《嫣然天使基金申请须知》的前提下,填写表格;2.申请资料报送中国红十字基金会嫣然天使基金项目管理办公室。
?第五届“BAZAAR明星慈善夜”创始人李亚鹏获十大慈善明星奖之最大奖项——“特别爱心奖”(未领)嫣然天使基金标志?“AAC2007艺术中国·年度影响力”颁奖盛典获“年度艺术慈善奖”?慈善(南方)盛典”创始人李亚鹏获“慈善明星奖”(之前他一直拒绝领奖,这是他第一次拿慈善奖,“因为现在做慈善已经不是我个人的行为,而是我们整个团队都在做这件事情,我想拿奖对大家应该是个精神上的鼓励。”) ?“2009北京印·时尚大典”创始人李亚鹏及阿里之行医疗队获“时尚档案慈善事业”大奖。 ?“2009年度中华慈善奖颁奖典礼”,嫣然天使基金获“最具影响力慈善项目”奖项。
关于设立投资担保公司的说明书
目录 一、项目概述 (3) 二、设立投资担保公司的价值 (4) 三、投资担保策略 (7) 四、投资担保公司管理及架构 (9) 五、投资担保公司融资担保流程 (10) 六、投资担保公司收益及分配 (13) 七、其他中介机构 (16)
一、项目概述 (一)项目名称:投资担保公司 (二)公司形式:有限责任公司 (三)注册资本:1亿元人民币。 (四)公司宗旨:为地方中小企业贷款提供担保,打通中小企业融资瓶颈,支持中小企业自主创新,完善创业发 展环境,拉动区域经济增长。 (五)出资人:----创业投资及战略合作伙伴。 (六)存续期限:10年 (七)担保对象:当地具有区域特色、高科技、创新型、农业产业化、能源类、循环经济模式、新经济模式等有 良好发展前景的企业。 (八)运作模式:同政策性银行、商业银行合作,运用担保的金融杠杆作用,为当地符合公司担保对象条件的企 业在银行借款提供担保,并为企业制定融资计划,进 行商业谈判然后签署债转股协议,取得被担保企业的 股份,最后通过良好的退出机制来获得回报。(九)目标区域:、株洲、等地区。
二、设立投资担保公司的价值 担保业在我国是一个新兴行业,其运营和监管一直处于摸索阶段,国家尚没有对行业进行整体的立法和规。监管主体不明确,缺乏统一的行业管理规,准入监管、日常运营监管和退出监管等监管容尚需完善。 2006年8月,银监会下发《关于银行业金融机构与担保机构开展合作风险提示的通知》,要求商业银行与投资担保公司开展业务合作时,要严格审查其资质条件,要求投资担保公司注册资本金必须在1亿元以上,必须是实缴资本,并且将逐步淘汰管理不规、运作模式不科学的担保机构。同时,国家正在积极设立担保业准入制度,构建担保业准入体系,并将担保业纳入相关部门的监管体系,担保业门槛将会进一步提高。 随着我国经济的发展、市场的进一步开放,投资担保公司将在日后的规过程中逐渐成为更加重要的非银行金融机构(将纳入监管围),担保业务的创新将会进一步扩大,发展的空间也是巨大的。现在正是我们进入担保业的良好时机,也是最后的机会。 (一)投资担保公司的作用
××××××担保有限公司公司经营发展战略和规划 ××××××担保有限公司在经营活动中,将以《中华人民共和国担保法》和最高人民法院《关于适用<中华人民共和国担保法>若干问题的解释》等法律法规为依据,严格遵循国家七部委《融资性担保公司管理暂行办法》和省、市相关规定的要求,以合法性、安全性、效益性和服务性为基本准则,坚持市场化运作、制度化管理、规范化操作,秉承防范与化解风险、诚实守信的经营原则,充分发挥自身独特的功能与机制设计上的优势开展相关经营活动。为此,特提出如下经营战略和规划: 一、公司经营范围与业务运作模式界定 1、主要担保对象 公司将以具有充分履约能力的企业和个人为主要担保对象,为其提供融资担保服务。 2、主要业务品种 根据我们对当前担保市场的调查和分析,确定公司的主要业务品种为: (1)中小企业贷款融资担保; (2)工程履约担保、诉讼财产保全担保、保函担保; (3)个人经营性贷款融资担保、消费、创业、出国留学等贷款担保; (4)二手房产交易担保、房产抵押贷款担保、房屋按揭、转按
揭贷款担保。 (5)担保配套服务。担保配套服务是指围绕担保项目由本担保公司提供的一系列中介服务。 3、主要担保模式设计 根据公司业务品种的需要,初步设计主要按以下几种担保模式开展担保业务。 (1)增信倍增贷款担保模式,适应于已达到银行授信准入条件的科技成长型企业,通过追加本公司的担保增信,倍增其贷款规模,增强银行贷款风险控制强度。 (2)循环额度贷款担保,一次性签订额度贷款合同,在规定的期限和额度内,可多次借款、逐笔归还、循环使用,适用于业务多元化企业。方便灵活、财务费用较低。 (3)到期一次性还款的贷款担保模式,其资金使用价值较好,但还款压力较大。 (4)在借款期内分期还款的贷款担保模式,包括均匀分期还款和个性化分期还款,资金成本较低,还款压力较小。 (5)临时过桥贷款担保模式,客户在短期内确定有对应的一笔现金流入,过程中又有物权作保证,担保人参与贷款资金运作和回笼全过程以控制风险。 (6)以应收账款质押为主的贷款担保模式,客户以短期优质的应收帐款质押作为主要反担保,申请本公司提供连带责任担保的银行贷款,加快资金周转,减少资金积压。
证券代码:002605 证券简称:姚记科技公告编号:2020-076 上海姚记科技股份有限公司 关于公司及子公司向相关银行申请综合授信额度 并为其提供担保的议案 本公司及董事会全体成员保证信息披露内容的真实、准确和完整,没有虚假记载、误导性陈述或重大遗漏。 上海姚记科技股份有限公司(以下简称“公司”)于2020年6月4日召开第五届董事会第二次会议审议通过了《关于公司及子公司向相关银行申请综合授信额度并为其提供担保的议案》,现将相关内容公告如下: 一、概述 公司及控股子公司浙江万盛达扑克有限公司(以下简称“万盛达”)为满足其运营发展周转所需流动资金,拟向相关银行申请合计28,000万元的综合授信额度(含前次未到期的授信和担保额度),包括流动资金贷款、银行承兑汇票、 公司及全资子公司启东姚记扑克实业有限公司(以下简称“启东姚记”)、公司实际控制人姚文琛先生和邱金兰女士为支持公司及控股子公司万盛达的经营发展,拟为公司及万盛达公司向上表中相关银行申请的综合授信额度提供担保,保证期间为主合同项下债务人每次使用授信额度而发生的债务履行期限届满之日起两年。上述担保中,公司实际控制人姚文琛先生和邱金兰女士不向公司收取任何担保费用,也不需要公司提供反担保。 根据《深圳证券交易所创业板股票上市规则》、《公司章程》等相关规定,上
述担保事项在董事会审批权限范围内,不需提交公司股东大会审议,本次担保不构成关联交易。 二、被担保人基本情况 1、上海姚记科技股份有限公司 上海姚记科技股份有限公司成立于 1989 年 09 月 13 日,注册地址为上海市嘉定区黄渡镇曹安路4218号,法定代表人为姚朔斌,注册资本为 39,820.9187 万元人民币,公司信用状况良好。主要经营范围:从事计算机软硬件技术领域的技术研发、技术咨询、技术服务、技术转让,扑克牌,包装装潢印刷,零件印刷,经营本企业自产产品及技术的出口业务,经营本企业生产、科研所需原辅料、机械设备、仪器仪表、零配件及技术的进出口业务(国家限定公司经营和国家禁止进出口的商品及技术除外),经营进料加工及“三来一补”业务,设计、制作各类广告,利用自有媒体发布广告,自有房屋租赁,投资管理,咨询服务,经营性互联网文化信息服务。【依法须经批准的项目,经相关部门批准后方可开展经营活动】 主要财务数据: 单位:元 2、浙江万盛达扑克有限公司 浙江万盛达扑克有限公司成立于 2004 年 5 月 24 日,注册地址为浙江省兰溪市江南高新工业园区,法定代表人为盛永兴,注册资本为 6667 万元人民币,主要经营范围:扑克、玩具、纸制品制造、销售,广告设计制作,包装装潢、其他印刷品印刷;货物及技术进出口业务。万盛达公司为公司控股子公司,信用状况良好。 主要财务数据: 单位:元
科技园区投融资案例借鉴分析 (一)投融资改革中的“中关村模式” 简单说来,“中关村模式”就是针对企业发展的不同阶段制定不同的投融资促进模式和方案。 详细地来说,这种具有中关村特色的投融资模式充分发挥政府的引导作用,根据高新技术产业发展的规律,针对不同发展阶段的企业不同融资需求特征,以企业信用建设为基础,充分利用市场机制,采用多方合作的方式,既拓宽间接融资渠道,又通过促进创业投资、组织发行高新技术企业集合信托计划和企业债券、推动企业改制上市等多渠道,来提高直接融资比重,制定了促进不同阶段企业发展的投融资政策体系,把各种资源,包括政策、资本、技术、市场整和在一起,支持企业,然后功成身退。 中关村科技园区针对处于初创期、快速成长期和稳定发展期的企业制订了有针对性的投融资促进方案。针对初创期企业融资难的问题,中关村管委会为填补“投资空白”,解决市场失灵,在全国率先设立了创业投资引导资金,实施创业投资企业风险补贴政策,研究促进天使投资发展,搭建了创业投资促进工作平台;针对快速成长期企业担保难、贷款难问题,中关村管委会设立了针对重点企业群体的担保贷款绿色通道、组织发行高新技术企业集合信托计划和企业债券、开展信用贷款试点,搭建了专门服务于中小型高新技术企业的贷款担保平台;在促进稳定发展期企业发展方面,主要是实施改制上市资助政策,促进具有一定规模企业的产(股)权流动,使企业借助资本市场进一步做强做大,积极开展中关村企业股份报价转让试点,搭建产(股)权交易平台和专业化的企业改制上市服务平台。 1、针对初创企业的创投促进体系 针对初创期企业出现的“投资空白”,中关村管委会创新了财政资金的使用方式,建立投资引导放大机制,促进创业投资在园区的业务聚集和物理聚集,于2001年年底设立了“中关村科技园区创业投资引导资金”,对经认定的创业投资机构投资于园区企业给予一定比例的跟进投资资金支持。政府虽然以股权方式出资,但不介入投资管理,而是委托专业投资管理机构进行管理,采用市场化的运作机制。这样既能实现政府的公共政策目标,又可以大大
由于银行审核严格,需要资金周转的人士都会找上贷款担保公司进行融资,俗话说“货比三家”,那么在深圳担保公司中哪家更好呢?更加专业呢? 一、政府特批成立的担保公司,很多人以为担保公司只有民营的,正是因为担保市场鱼龙混杂,特别成立一批担保公司用来规范不正当竞争市场,深圳较为有名的担保公司如下: 1、深圳市中小企业信用融资担保集团有限公司 深圳市中小企业信用融资担保集团有限公司的前身是深圳市中小企业信用担保中心,成立于1999年12月28日,是深圳市政府成立的专业担保机构,现注册资本18亿元人民币,担保机构资信评级AAA,资本市场评级AA+级。 股东:深圳市投资控股有限公司、中国东方资产管理公司。 2、深圳市不动产融资担保股份有限公司 深圳市不动产融资担保股份有限公司是根据国家建设部和中国人民银行《住房置业担保管理试行办法》(建住房〔2000〕108号文)的有关规定,经深圳市国土资源和房产管理局审核,深圳市人民政府批准《关于以发起方式设立深圳市不动产担保股份有限公司的批复》(深府股[2002]43号文)后,于2002年12月20日由深圳市土地房产交易中心等发起人发起设立的专业不动产担保机构。 公司股东由深圳市中小企业信用融资担保集团有限公司、深圳市规划国土房产信息中心和卓越集团(实际控制人)旗下两家公司等组成。 二、不只是政府特批,深圳担保行业之所以居于全国领先地位,其开放的市场孕育了广大民营担保公司如雨后春笋般的崛起: 1、深圳市高新投集团有限公司 深圳市高新投集团有限公司(以下简称“高新投”)成立于1994年12月,是深圳市政府为解决中小科技企业融资难问题而设立的担保机构,也是国内最早设立的专业担保机构之一。集团注册资本48.52亿元,股东为深圳市投资控股有限公司、深圳远致富海三号投资企业(有限合伙)、深圳市财政金融服务中心、恒大企业集团有限公司、深圳市远致投资有限公司、深圳市海能达投资有限公司、深圳市中小企业服务署。高新投核心业务为:融资与金融产品担保、保证担保、资产管理。 2、深圳深业融资担保有限公司 深圳深业融资担保有限公司是经深圳市贸工局批准,于2005年1月成立的中外合资专业投资担保公司,主要为深圳市中小企业和个人提供担保服务及相关信息咨询服务。 3、深圳共盈通金融服务有限公司