
生长素在根部运输的几种方式
茎中生长素的运输方向是形态学上端到下端的极性运输,那么,在植物根中生长素的运输方向是什么呢?
按照目前流行的各种高中生物教辅资料中,在解释把植物横放后根的向重力性时生长原因时,一般都认为:是根尖产生了生长素,在重力的作用下,由根尖的远地侧向近地侧横向运输,然后再向根基部方向运输,由于根对生长素很敏感,故抑制了近地侧根细胞的生长,使远地侧根细胞生长速度较快而出现向重力性生长。也就是说,根中生长素是由根尖向根基部的极性运输。然而,这种说法是不正确的。
1.根中生长素的来源
IAA主要是在植物的顶端分生组织中合成的,合成后即被运输到植株各部和根中。IAA的运输是单方向的,只能从植物顶端往下运输而不能反过来从下面往顶端运输,所以称为极性运输。根据根向重力性Evans-Moore模型,根直立生长时茎尖运向根冠的IAA在根中四周平均分布。高中教材上也提到,生长素主要产生于幼嫩的芽、叶,发育中的种子等。生长素在植物中分布很广,但以生长旺盛的器官和组织如正在生长的茎尖和根尖的分生组织、胚芽鞘、受精后的子房和幼嫩的种子等含量较多,而在衰老的组织和器官中含量甚少。从这里可以这样理解:根尖尤其是根尖分生组织中的较多生长素不是根尖产生的,而是茎尖产生后向下运输一直运输到根尖,并作用于根尖的。
2.根中生长素的极性运输
植物根中的生长素也表现极性运输的性质,不过是由根基部向根尖方向的运输,即向顶运输。
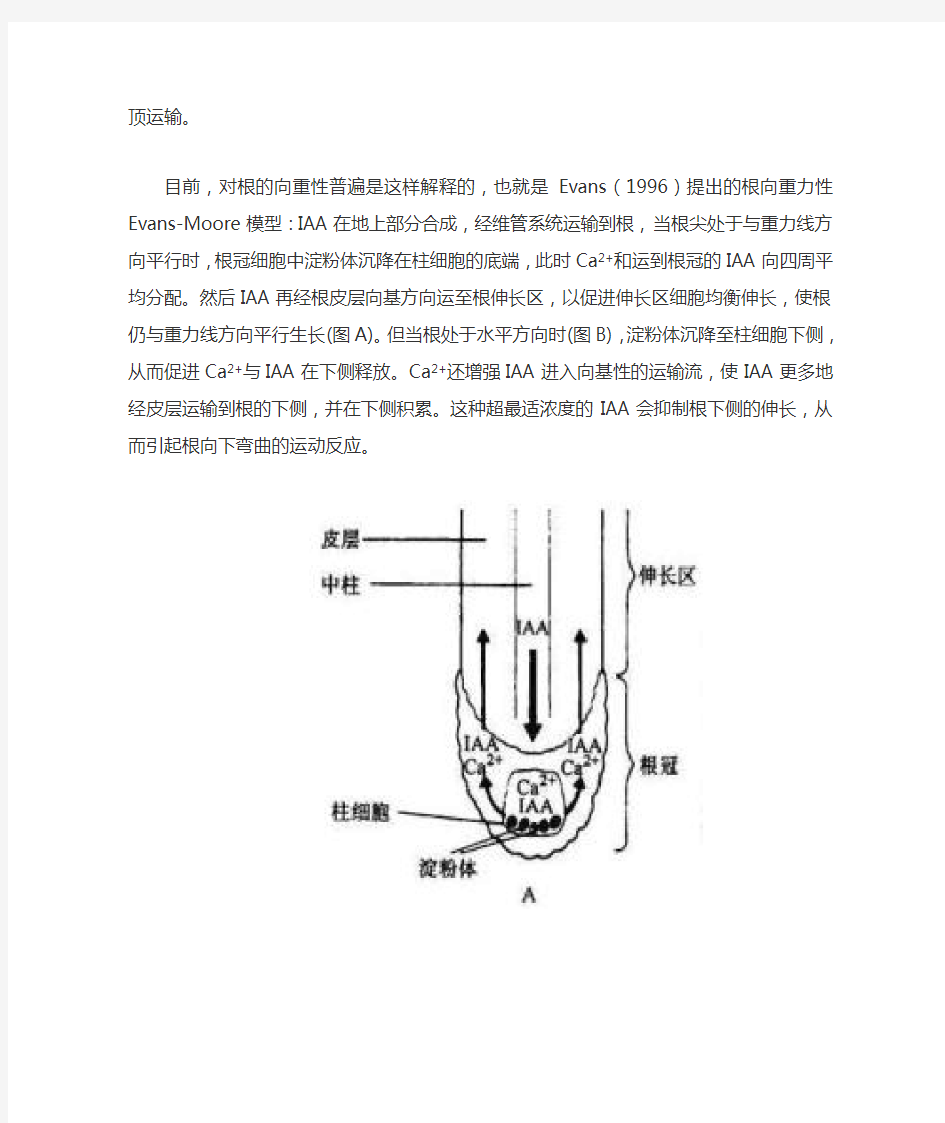
目前,对根的向重性普遍是这样解释的,也就是Evans(1996)提出的根向重力性Evans-Moore模型:IAA在地上部分合成,经维管系统运输到根,当根尖处于与重力线方向平行时,根冠细胞中淀粉体沉降在柱细胞的底端,此时Ca2+和运到根冠的IAA向四周平均分配。然后IAA再经根皮层向基方向运至根伸长区,以促进伸长区细胞均衡伸长,使根仍与重力线方向平行生长(图A)。但当根处于水平方向时(图B),淀粉体沉降至柱细胞下侧,从而促进Ca2+与IAA在下侧释放。Ca2+还增强IAA进入向基性的运输流,使IAA更多地经皮层运输到根的下侧,并在下侧积累。这种超最适浓度的IAA会抑制根下侧的伸长,从而引起根向下弯曲的运动反应。
根据以上可知,根中生长素在根的微管系统中是从根基部向根尖的极性运输。
但是有人用14C-IAA体外供给植物的试验发现,根部吸收的大多随蒸腾流沿导管上运并转至其他部位。这是怎么回事呢?
实际上,除了极性运输之外,在植物体中还存在非极性的长距离运输方式。非极性运输的方向决定于两端有机物浓度差等因素,是通过韧皮部进行运输的。实际上植物体内的大部分束缚型生长素也是通过韧皮部运输的。束缚型生长素无生理活性,在植物体内的运输也没有极性,当束缚型生长素再度水解成游离型生长素时,又表现出生物活性和极性运输。也有人切除根尖或施用IAA的拮抗剂发现一般都能促进根的伸长,说明根尖中的IAA的含量通常处于抑制其生长的范围内。
所以,再结合前面对根的向重力性的解释可以看出,生长素在根(茎)的微管系统中是由根基部向根尖的极性运输,使根尖分布较多,由于根尖浓度大于靠近根尖部位,这种浓度差使得生长素又从根尖沿韧皮部向根基部方向进行非极性运输。而这时的生长素很可能就是束缚型生长素。这可能对侧根的发生具有重要意义。
生长素在植物的胚芽鞘尖端可以横向运输,是因为尖端没有维管束,当幼根处于水平方向时,根尖部位会发生横向运输吗?根据根向重力性Evans-Moore模型,当根从垂直方向转到水平方向时,根冠柱细胞中淀粉体向重力方向沉降,从而促进Ca2+和IAA在下侧释放。从这里可以看出,在根尖的根冠部位存在生长素的横向运输。
综上所述,根中生长素的运输方向有极性运输、非极性运输、横向运输。根中的生长素主要来自地上部分的茎的幼嫩部位,通过微管系统从根基部向根尖进行极性运输;根尖积累的生长素可以通过韧皮部从根尖向根基部进行非极性运输;根尖的根冠部位在外力作用下(重力)可以发生横向运输。根的形态学上端是根基部,形态学下端是根尖。
附:下面是关于生长素运输的最新研究进展节选
2014年浙江省高考题:
植物生长素的产生运输和作用 生长素(auxin)是一类含有一个不饱和芳香族环和一个乙酸侧链的内源激素,英文简称IAA,国际通用,是吲哚乙酸(IAA)。4-氯-IAA、5-羟-IAA、萘乙酸(NAA)、吲哚丁酸等为类生长素。1872年波兰园艺学家谢连斯基对根尖控制根伸长区生长作了研究;后来达尔文父子对?草胚芽鞘向光性进行了研究。1928年温特首次分离出这种引起胚芽鞘弯曲的化学信使物质,命名为生长素。1934年,凯格等确定它为吲哚乙酸,因而习惯上常把吲哚乙酸作为生长素的同义词。 生长素在扩展的幼嫩叶片和顶端分生组织中合成,通过韧皮部的长距离运输,自上而下地向基部积累。根部也能生产生长素,自下而上运输。植物体内的生长素是由色氨酸通过一系列中间产物而形成的。其主要途径是通过吲哚乙醛。吲哚乙醛可以由色氨酸先氧化脱氨成为吲哚丙酮酸后脱羧而成,也可以由色氨酸先脱羧成为色胺后氧化脱氨而形成。然后吲哚乙醛再氧化成吲哚乙酸。另一条可能的合成途径是色氨酸通过吲哚乙腈转变为吲哚乙酸。 在植物体内吲哚乙酸可与其它物质结合而失去活性,如与天冬氨酸结合为吲哚乙酰天冬氨酸,与肌醇结合成吲哚乙酸肌醇,与葡萄糖结合成葡萄糖苷,与蛋白质结合成吲哚乙酸-蛋白质络合物等。结合态吲哚乙酸常可占植物体内吲哚乙酸的50~90%,可能是生长素在植物组织中的一种储藏形式,它们经水解可以产生游离吲哚乙酸。 植物组织中普遍存在的吲哚乙酸氧化酶可将吲哚乙酸氧化分解。 生长素有多方面的生理效应,这与其浓度有关。低浓度时可以促进生长,高浓度时则会抑制生长,甚至使植物死亡,这种抑制作用与其能否诱导乙烯的形成有关。生长素的生理效应表现在两个层次上。 在细胞水平上,生长素可刺激形成层细胞分裂;刺激枝的细胞伸长、抑制根细胞生长;促进木质部、韧皮部细胞分化,促进插条发根、调节愈伤组织的形态建成。 在器官和整株水平上,生长素从幼苗到果实成熟都起作用。生长素控制幼苗中胚轴伸长的可逆性红光抑制;当吲哚乙酸转移至枝条下侧即产生枝条的向地性;当吲哚乙酸转移至枝条的背光侧即产生枝条的向光性;吲哚乙酸造成顶端优势;延缓叶片衰老;施于叶片的生长素抑制脱落,而施于离层近轴端的生长素促进脱落;生长素促进开花,诱导单性果实的发育,延迟果实成熟。 近年来提出激素受体的概念。激素受体是一个大分子细胞组分,能与相应的激素特异地结合,尔后发动一系列反应。吲哚乙酸与受体的复合物有两方面的效应:一是作用于膜蛋白,
第三章:植物的激素调节 第一节生长素的发现过程 向光性:在单侧光的照射下,植物朝向光源方向生长的现象叫做向光性。 一、生长素的发现过程 1.达尔文实验——胚芽鞘尖端是感受单侧光的部位,向光弯曲的部位在胚芽鞘尖端下部 1向光生长 2 不生长不弯曲3直立生长4向光弯曲 2.詹森的实验 图3-4詹森的实验示意图 影响可穿过琼脂由尖端向下传递 3.拜尔的实验——尖端产生刺激在下部分布不均引起弯曲生长 4.温特的实验——尖端产生的刺激是某种物质(温特将其命名为生长素) 5.化学本质:吲哚乙酸(IAA),还有苯乙酸(PAA),吲哚丁酸(IBA)等。 6.植物激素:(1)概念:由植物体内产生,能从产生部位运送到作用部位,对植物的生长 发育有显著影响的微量有机物,称作植物激素。
(2)其他植物激素:赤霉素,细胞分裂素,脱落酸,乙烯 二、生长素的产生、运输和分布 1.产生:部位:幼嫩组织,分生组织,幼嫩的芽、叶和发育中的种子,胚芽鞘尖端。 机理:色氨酸→生长素 2.运输:横向运输:发生在尖端部位 影响因素:光,重力,水等(与向性运动有关) 纵向运输极性运输:部位:发生在幼嫩部位如胚芽鞘、芽、茎尖、根尖等 方向:只能从形态学上端运输到形态学下端,不能反过 来运输,也就是只能单向运输 运输方式:为主动运输方式 影响因素:载体,ATP,O2 非极性运输:部位:发生在成熟组织中的韧皮部 方向:双向运输 3.分布: 部位:植物体的各个器官中都有分布,但相对集中在生长旺盛的部分 特点:生长旺盛部位 >衰老成熟部位 三、胚芽鞘向光弯曲生长的分析 单侧光→胚芽鞘尖端 尖端以下部位茎向光弯曲(有利于光合作用) ∣ 四、植物向性运动的人工实验方法归类 1. 暗盒开孔类(如下图) 直立生长向光弯曲 2. 云母片插入类 形态学上端 形态学下端形态学上端 形态学下端 3 4 生长素极性运输生长素分布不均 (内因) 光光 光 光 光
生长素在根部运输的几点认识 在人教版新课标高中生物必修3第3章第1节《植物生长素的发现》一节中,茎中生长素的运输方向是形态学上端到下端的极性运输,那么,在植物根中生长素的运输方向是什么呢? 按照目前流行的各种高中生物教辅资料中,在解释把植物横放后根的向重力性时生长原因时,一般都认为:是根尖产生了生长素,在重力的作用下,由根尖的远地侧向近地侧横向运输,然后再向根基部方向运输,由于根对生长素很敏感,故抑制了近地侧根细胞的生长,使远地侧根细胞生长速度较快而出现向重力性生长。也就是说,根中生长素是由根尖向根基部的极性运输。然而,这种说法是不正确的。 1根中生长素的来源 IAA主要是在植物的顶端分生组织中合成的,合成后即被运输到植株各部和根中。IAA的运输是单方向的,只能从植物顶端往下运输而不能反过来从下面往顶端运输,所以称为极性运输[1]。根据根向重力性Evans-Moore模型,根直立生长时茎尖运向根冠的IAA在根中四周平均分布[2]。高中教材上也提到,生长素主要产生于幼嫩的芽、叶,发育中的种子等。生长素在植物中分布很广,但以生长旺盛的器官和组织如正在生长的茎尖和根尖的分生组织、胚芽鞘、受精后的子房和幼嫩的种子等含量较多,而在衰老的组织和器官中含量甚少[2]。从这里可以这样理解:根尖尤其是根尖分生组织中的较多生长素不是根尖产生的,而是茎尖产生后向下运输一直运输到根尖,并作用于根尖的。 2根中生长素的极性运输 植物根中的生长素也表现极性运输的性质,不过是由根基部向根尖方向的运输,即向顶运输[2]。 目前,对根的向重性普遍是这样解释的,也就是Evans(1996)提出的根向重力性Evans-Moore 模型:IAA在地上部合成,经维管系统运输到根,当根尖处于与重力线方向平行时,根冠细胞中淀粉体沉降在柱细胞的底端,此时Ca2+和运到根冠的IAA向四周平均分配。然后IAA再经根皮层向基方向运至根伸长区,以促进伸长区细胞均衡伸长,使根仍与重力线方向平行生长(图A)。但当根处于水平方向时(图B),淀粉体沉降至柱细胞下侧,从而促进Ca2+与IAA在下侧释放。Ca2+还增强IAA 进入向基性的运输流,使IAA更多地经皮层运输到根的下侧,并在下侧积累。这种超最适浓度的IAA会抑制根下侧的伸长,从而引起根向下弯曲的运动反应[3]。
吲哚乙酸的运输详解 植物激素特点之一是可以广泛地在植物体内各组织和器官内移动,运输是一种天然的调节特定组织或器官内活性激素水平的手段。 游离态IAA具有极性运输的特点,在植物胚芽鞘、幼茎及幼根中,形态学上端的IAA 只能运向形态学下端,这种现象称为极性运输(po1ar transport)。IAA极性运输是由某些载体介导的主动运输。呼吸链抑制剂氰化物和解偶联剂DNP均能抑制IAA极性运输,说明IAA 极性运输需要有氧呼吸提供能量。另外,植物组织对放射性IAA的吸收受到非放射性IAA 的部分抑制,说明放射性标记与非标记的IAA竞争数量有限的载体位点。 Goldsmith(1977)提出了“化学渗透极性扩散假说(chemiosmotic polar diffusion hypothesis)” (图8-4,见e DH08-04)。该假说认为:位于某个细胞基部的IAA输出载体从细胞内单向输出IAA-,IAA-进入细胞壁空间后即被质子化为IAAH,IAAH扩散通过细胞膜,顺着其浓度梯度进入其下部相邻的细胞内。由于细胞质的pH约为7.2,IAAH进入到细胞质后,几乎所有的都被解离成阴离子IAA-的形式。IAA-不能扩散通过细胞膜,只能依靠输出载体输送至细胞壁空间。按此规律,IAA顺序通过纵向排列的细胞柱向形态学下端运输。在此过程中,位于质膜上的H+-ATPase不断地将H+从胞内泵出,以防止H+在胞内积累,并维持细胞壁酸性环境和适宜的跨膜电势梯度,以提供能量。 现已清楚,IAA-也可通过载体介导的2H+-IAA-同向共运输体进入,质膜上一个小的AUX1/LAX透酶家族是生长素的输入载体,可同时转运2个质子和IAA-同向进入细胞质。PIN家族是IAA的输出载体。 在拟南芥中,存在8个基因编码PIN蛋白,研究表明PIN1、PIN2 、PIN3 、PIN4 和PIN7 都参与了生长素极性运输,它们介导不同组织的生长素输出。PIN1是发现最早的,也是主要负责生长素的从茎顶端向根尖的极性运输。最近几年研究者还发现另外一种依赖ATP 的转运蛋白参与生长素的极性运输,它们属于多种药物抗性/ 磷酸糖蛋白家族(multidrug resistance/ P-glycoprotein,MDR/ PGP),或ATP 结合盒(ATP-binding cassette ,ABC) 转运蛋白超级家族中的“B”亚家族(ABCB)。ABCB基因发生缺陷的拟南芥和玉米植株均表现为不同程度的矮小,改变向地性,并且生长素的输出减少。它们均匀地分布在质膜上,驱动生长素的依赖ATP的输出,PINs协同ABCBs调控生长素的极性运输。 某些化合物能够专一地抑制IAA极性运输,如2,3,5-三碘苯甲酸(TIBA, 2,3,5-triiodobenzoic acid)、9-羟基氟-9-羧酸(HFCA, 9-hydroxyfluorine-9-carboxylic acid, 又叫形态素)和N-1-萘基邻氨甲酰苯甲酸(NPA, N-1-naphthylphthalamic acid)。TIBA及NPA通过阻止生长素外流而阻止生长素的极性运输。它们的抑制作用均为非竞争性的,说明它们与IAA在输出载体上占有不同的位置,它们与载体的结合引起载体蛋白构型发生改变,抑制载体对IAA输出。
试析生长素的运输与茎的向光性 南海实验学校高中部游云霞 关键词:生长素、横向、向光性。 摘要:本文综述了植物向光性与生长素运输方式的关系。横向运输和极性运输的作用机理和影响横向运输的因素,以及生长素如何引起细胞伸长。 正文: 植物的一生离不开激素,各种激素相互协调共同对植物的生命活动起着重要的调节作用,植物之所以表现出生长现象,那是因为生长素在促进其生长方面占主导地位。在教学中我们常要分析这样一些问题,即对燕麦胚芽鞘作各种处理,培养一段时间后,判断胚芽鞘生长情况(如图所示) 锡箔片琼脂(说明:⑵在胚芽鞘尖端下用锡箔片遮 光⑶切去胚芽鞘的尖端但在切口上放一含有一定浓度的生长素琼脂小块。⑷如⑶处理后放在暗盒里。以上都给予单侧光)结果:⑴⑵弯向光源生长,⑶⑷直立生长。? 那么横向运输为什么会引起生长素分布不均?下面就从生长素的运输方面来探讨一下这个问题。 一、横向运输 1. 作用机理:所谓横向运输就是指生长素由茎(光源)的一侧横向移动到另一侧(背光)的运输方式.由此造成的生长素分布不均实际上应与电荷分布有关.我们知道,生长素即吲哚乙酸(IAA)是带弱酸性的,在细胞中常以阴离子( IAA-)形式存在,而对植物来说,单方向的光照会引起器官尖端不同部位产生电势差,向光一侧带负电荷,背光一侧带正电荷,这样一来,生长素带弱酸性的阴离子则向带正电荷的背光一边移动,再向下运输,从而引起尖端下背光一侧生长素分布多,细胞纵向伸长快,向光一侧分布少,细胞纵向伸长慢,使植物弯向光源生长. 那么植物在怎样的条件下会发生横向运输? 2.影响横向运输的因素. ①单侧光:生长素发生横向运输引起向光性与单侧光密切相关. 为什么单侧光会引起横向运输?主要是由于植物的受光不均.假如将照射植物的单侧光改成直射光使之受光均匀或不照光,则植物不出现向光性生长.这是因为在这种情况下,电荷分布均匀,此时生长素只进行极性运输.而不表现横向运输,说明单侧光是引起生长素横向运输的原因之一。 ②尖端:横向运输不可缺少的部位. 文章开头提到的图中;实验证明(1)(2)(3)都具有生长素.且都有单侧光照射.但(1)(2)弯向光源生长而(3)却直立生长.是什么造成(3)与(1)(2)的不同呢?是尖端即(3)没有尖端部位,也就是说横向运输只发生在尖端部位.是否如此?通过下面一组实验就知道了,即将胚芽鞘作如图处理: (锡箔片) (1) (2) (3) (4)
生长素 29.(16分) 向光性是高等植物中广泛存在的生理现象,是植物适应环境变化的一种体现。研究表明,单侧光照射下水稻的根会发生背光弯曲即“负向光性”。为研究IAA对水稻根负向光性运动的影响及有关作用机理,研究人员进行了相关的实验。 (1)已知Ca2+作为信号分子,在植物的多种信号转导及生长发育过程中起着重要的作用。为探究Ca2+是否会影响稻根中IAA的分布,研究人员用加入H2O、CaCl2溶液、LaCl2溶液(Ca2+通道阻断剂)以及后两者混合液的四组培养液分别培养水稻秧苗刚长出的根,在单侧光的照射24h后,四组稻根均出现负向光性,每组根中IAA的分布结果如下图。 由结果可知,在单侧光照下对照组中IAA的分布情况是,而Ca2+作为信号分子。与对照组相比,后两组实验的结果表明Ca2+能,从而进一步证明Ca2+在根负向光性运动中对IAA 分布的影响。 (2)目前已知cpt1基因编码的CPT1蛋白是水稻胚芽鞘向光性运动过程中IAA横向运输的重要载体。为探究cpt1基因是否与水稻根负向光性运动有关,研究人员对水稻秧苗刚长出的根分别进行不同处理,24h后测量稻根弯曲度(处理条件及结果见下表)。同时研究人员还测定了各组稻根中cpt1基因表达量(结果如下图)。 上图中的1~4表示黑暗条件下cpt1基因的表达量,7表示H2O处理组的表达量,5、6、8应分别是处理的结果。由此可知,外源施加的四种试剂对稻根中cpt1 基因表达量的影响与它们对稻根弯曲度的影响是一致的。在此实验结果基础上,并结合科研人员测定的单侧光照下cpt1 基因缺失突变体水稻根的向光侧和背光侧IAA均匀分布这一事实,推测CPT1蛋白在根中也是。 (3)综合上述实验结果推测,在Ca2+信号作用下,单侧光照射下水稻根内的IAA通过 ,导致IAA在向光侧与背光侧分布不均匀;由于根对IAA浓度,使得两侧的生长速度表现为,
(一)花及花序的分化与形成:烤烟生长到一定时期,大约在团棵后不久,就开始从营养生长转向生殖生长。首先是茎生长点分化的转变,在营养生长时,茎的生长点不断分化,形成新的叶子,结果使主茎伸长,叶片增多,当转入生殖生长期时,茎部的生长点则分化成生殖器官。其次是分枝式的转变,处在营养生长时期的茎,是按单轴式分枝,但当植株发育到生殖生长时,则按合轴式分枝,茎顶端分化为花,顶花下方的腋芽和副芽,则发展成为分枝,这种现象在植株外部形态上,还不能马上表现出来,直到全部叶片展开,主茎顶部的花发育成花蕾,生殖生长才明显表现出来。(二)花序分化的特征:在营养生长时,茎的生长点是一个平面体,周围有许多分化程度不同的幼叶,当转入生殖生长时,平面体突起而呈尖头状,约有10片左右的幼叶环抱着生长点,使顶芽变得饱满起来,此时主茎叶片数已经固定,生长点分化出3~4个小突起,这就是第一朵花和3个顶生花的原基。(三)烟草花序的发展:烟草花序是有限的聚伞形花序。烟草转入生殖生长时,顶芽发育成为第一朵花,而腋芽和副芽则变成了花枝,同时2个或3个花枝与第一朵花分布在一个平面上,形成三角形向各个方向发展。如此循环分化,花愈开愈多,花序也愈来愈复杂,同时主茎顶端也愈来愈开展,终于形成一个圆锥形花丛。 其他植物激素对植物生命活动的调节一、知识结构赤霉素细胞分裂素其他植物激的种类和作用乙烯其他植物激素脱落酸植物生长调节剂的应用二、教学目标1、列举其他植物激素。2、评述植物生长调节剂的应用。3、尝试利用多种媒体,搜集并分析植物激素和植物生长调节剂的资料。三、教学重点、难点及解决方法1、教学重点及解决方法[教学重点]其他植物激素的种类和作用。[解决方法]利用教材中的图,让学生进行图表转换,列出植物激素的种类和作用。2、教学难点及解决方法[教学难点]植物生长调节剂的应用。[解决方法]利用教材中的资料分析,让学生开展说一说活动,引导学生深入认识生物科学和技术的应用。四、课时安排1课时。五、教学方法直观教学法。六、教具准备图片资料。七、学生活动学生分析图文资料,得出结论。八、教学程序(一)明确目标(二)重点、难点的学习与目标完成过程[问题探讨]呈现教材P53木瓜催熟柿子的图,回答课本中的讨论题。学生阅读教材P53相关内容,了解其他植物激素的存在。呈现教材P54图,让学生针对图解中的事实,进行图表转换,列出植物激素的种类和作用表。激素名称主要合成部位主要作用赤霉素细胞分裂素脱落酸乙烯各种植物激素生理功能不同,但从生理作用上可分为二类:一类是具有明显促进生长发育的功效。如生长素、赤霉素、细胞分裂素;另一类则主要抑制生长发育,如脱落酸。植物的个体发育受多种激素的调节,不同时期有不同的主要激素起主导的调节作用。多种激素相互协调,共同完成对植物生命活动的调节。例如科学家在对黄化豌豆幼苗切段的研究中发现:生长素促进植物生长主要在于促进细胞纵向伸长,随着生长素浓度不断增高,这种促进细胞纵向生长的功效越强。但当生长素浓度增高到一定程度时,就会促进乙烯的合成。乙烯一方面抑制细胞纵向伸长,
生长素的极性运输 林科院 9310031 傅建平 摘要生长素的极性运输与植物生长发育密切相关并受许多因素调控,研究植物生长素的极性运输具有十分重要的意义。本文综述了生长素极性运输的研究进展情况。 关键词生长素极性运输抑制剂输出载体输入载体 生长素是第一个被发现的植物激素。生长素在植物叶原基、幼叶以及发育的种子等部位合成,沿茎或根进行传输。生长素在茎切段试验和胚芽鞘试验中表现刺激细胞伸长生长,它的主要生理作用还包括诱导不定根发生促进维管束分化、控制向性生长和顶端优势,以及调节植物开花坐果等生理过程。在植物茎尖合成的生长素需要通过极性运输到达靶细胞才能调节植物的生长和发育, 因此生长素极性运输广泛参与植物的叶片发育、花的分化、维管的分化、胚胎发育、光形态建成以及侧根的发育等。。早期对生长素极性运输的研究主要采取施加极性运输抑制剂如NPA、HFCA、TIBA、CA的方法。后来通过对生长素极性运输突变体的研究, 从分子水平上为生长素极性运输的化学渗透偶联学说提供了新的证据, 进一步发现了极性运输与植物的生长发育现象之间的联系。 1生长素的极性运输特点 生长素的极性运输是指生长素在植物体内由形态学的上端向形态学的下端单向运输的现象。在高等植物茎中, 生长素由茎尖合成位点极性运输到茎基部的作用位点。生长素极性运输具有依赖于能量、需要O2、对温度敏感、随年龄增加而减弱等生理特点,是一种需要消耗代谢提供能量的主动运输。其运输的速度比维管系统中的运输速度慢得多。极性运输需要消耗能量, 可逆浓度梯度运输。一些化合物如TIBA和NPA等能抑制生长素极性运输, 缺氧会严重地阻碍生长素的运输。此外, 生长素又有自动抑制现象, 即先发育的器官通过其合成并向外输出的生长素抑制后发育期器官生长素的输出。 2 极性运输处于主导地位 从整体组织看, 生长素的移动方向是向基性的。极性运输与能量消耗有关, 它包括生长素从上部细胞的透出和向下部细胞的透入, 透入时几乎不受代谢抑制剂或缺氧等因素的影响, 而透出却明显地受这些因素阻碍。显然, 极性运输是一种同代谢过程密切联系的主动运输。生长素的极性运输和横向运输是两个相对独立的运输过程, 前者与组织极性有关, 后者由重力作用引起, 它们在不同的情况下分别起作用并引起多种生理效应。那末, 在倒置的茎中, 是否会由于重力的作用而使生长素向形态学的顶端逆极性运输呢?当然, 很难设想在这情况下感应重力的横向运输机制完全不影响生长素的运输, 但正如实验所表明的那样, 茎的倒置并不能改变生长素的总的运输方向, 也就是说, 极性运输仍处于主导地位。 3抑制剂对生长素极性运输的调控 生长素输出载体是极性运输的主要调控部位。目前对生长素极性运输调控的研究主要集中在NPA结合蛋白对输出蛋白活性的调节上。常用抑制剂三碘苯甲酸( TIBA )和萘基邻氨甲酰苯甲酸(NPA)研究生长素极性运输, 它们对输出蛋白起负调控作用: 与负责IAA运输的载体蛋白结合, 通过改变载体蛋白的构象, 抑制IAA 与载体蛋白的特异性结合, 从而抑制IAA的极性运输。
植物生长素的发现专题练习 1.在如图所示的实验中,对实验结果预测正确的是(甲~丙是在切去尖端后进行的处理 A.上述四组实验中最先发育为侧枝的是侧芽4 B.上述四组实验中最先发育为侧枝的是侧芽3 C.主枝仍能直立生长的只有丙 D.能弯向光源生长的只有丁 【答案】A 【解析】上述四组实验中最先发育为侧枝的是侧芽2,A、B错误;主枝仍能直立生长的是甲、丙,C错误;丁给以单侧光照,植物会弯向光源生长,甲、乙、丙不具有尖端,不能接受单侧光照,因此甲、乙、丙不能向光弯曲,D正确。 2.根据所学知识判断,下列说法正确的是(注:为不透光的锡箔帽和套) A.若探究苗尖端感光部位,应设置④⑤进行对照 B.若探究植物产生向光性的外因,应设置②③进行对照 C.若探究植物产生向光性的内因,应设置①⑥进行对照 D.图中弯曲生长的是①③⑤ 【答案】A 3.侧芽生长素的浓度总是高于顶芽,但是顶芽产生的生长素仍大量积存于侧芽部位,这是因为生长素运输的方式属于
A.主动运输B.极性运输 C.自由扩散D.渗透作用 【答案】A 【解析】根据题意分析可知:侧芽生长素的浓度总是高于顶芽,但是顶芽产生的生长素仍大量积存于侧芽部位,说明生长素是由低浓度到高浓度的运输,这种运输方式属于主动运输,故A项正确,B、C、D项错误。 4.下图为达尔文进行的某植物向光性研究的示意图,下列表述正确的是 A.该实验结果说明尖端是该植物的感光部位 B.该实验研究的目的是探讨植物是否具有向光性 C.省略第⑤组实验,该实验仍可获得确切的实验结论 D.实验处理一段时间后,②、③的尖端以下部位生长高度相同 【答案】A 【解析】据图分析,①②实验说明尖端是该植物的感光部位,A正确;该实验研究的目的是探讨植物感受光刺激的部位和发生弯曲的部位,B错误;省略第⑤组实验,则该实验不能获得确切的实验结论,C错误;实验处理一段时间后,②由于没有尖端,不能生长,而③直立生长,D错误。[来源:学,科,网Z,X,X,K] 5.某小组利用同一种植物幼苗进行如下实验,几天后,预测幼苗的茎仍直立生长的是 A.ⅠB.Ⅰ和Ⅱ C.Ⅱ和ⅢD.Ⅰ、Ⅱ和Ⅲ 【答案】A 【解析】分析图示可知,装置Ⅰ中幼苗处于暗室中,没有单侧光的刺激,因此幼苗将直立生长;装置Ⅱ的右侧上方有单侧光刺激,此时单侧光使生长素由向光一侧朝背光一侧运输,导致背光一侧生长素浓度高于向光一侧,背光一侧生长快,从而弯向光源生长;装置Ⅲ的右上方和右下方均有单侧光,因此与装置Ⅱ相同,茎表现为弯向光源生长;而根对生长素浓度最敏感,背光侧生长素浓度高反而抑制生长,因此向光一侧生长快,使根背光
植物生长素的运输特性及在植物生长中的调节作用辨析 李珊珊李建宏*(南京师范大学生命科学学院210046)李其柱(江苏省连云港市教育局教研室222006) 摘要本文对植物生长素的运输、生长素生物效应的两重性以及生长素对植物向地性的影响等相关问题进行简要介绍和辨析。关键词生长素极性运输两重性向地性 1植物生长素的运输问题 在植物体中,游离生长素的运输方式有两种:一种是和其他同化产物一样,通过韧皮部运输,其运输方向由两端有机物浓度差等多种因素决定,如成熟叶片中合成的吲哚乙酸在韧皮部的运输;另一种是仅局限于胚芽鞘、幼茎、幼根的薄壁细胞之间短距离单方向的极性运输[1]。 植物生长素的“极性运输”是其合成运输的一个重要特性。“从形态学的上端向形态学的下端”这一过程的成立是建立在“形态已形成”的基础上的。即必须是先确立了形态的极性,才明确运输的方向。形态学的上端即“远基端”(远离基部),下端为“近基端”(靠近基部)。极性运输过程是逆浓度的主动运输过程。产生极性运输的机理是细胞的下端有更多的离子态的吲哚乙酸(IAA)载体,而运出细胞膜外的IAA在处于低pH环境下变为分子状态,可进入下一个细胞,产生从上往下的运输链[2]。 对于“愈伤组织中生长素的运输是不是极性运输”的问题,一方面,这一问题已超出极性运输这一自然生物学现象的讨论范畴;另一方面,在一般的组织培养研究中,培养基中的生长素(或生长素类生长调节剂)浓度远高于植物体中的一般浓度,扩散进组织块的生长素的作用占绝对优势,讨论植物细胞间生长素的运输没有意义。从推断意义上看,由于愈伤组织尚未进行分化,没形成形态学的上端和下端,其生长素的运输不是极性运输。 燕麦胚芽鞘(注意:不是幼叶)的向光弯曲实验,揭示了生长素在胚芽鞘尖端合成,并且导致弯曲向光生长。虽然早期的实验认为,光照影响了生长素在胚芽鞘两侧的分布,导致背光面浓度增加,生长加快,但后来更精细的测定结果并不支持这一结论,在其他植物中也未发现这样的现象。因此,向光弯曲生长是否与光诱导生长素分布不均有关尚无更多的实验证实[2]。这一未定论的知识点在中学教学中不宜作为定论来强调。 关于“生长素从切割下来的胚芽鞘向琼脂块当中的运输是何种运输方式”也是一个容易混淆的问题。实际上生长素从切割下来的胚芽鞘界面向琼脂块中的运动(不是运输)是化学扩散作用,与胚芽鞘的生理活动无关。生长素向琼脂块中扩散的速度,主要取决于两者的浓度差。但胚芽鞘切口处的生长素浓度却是极性运输的结果。因此,总的来看,在胚芽鞘中生长素通过极性运输积累在下端,在下端的切口处通过化学扩散进入琼脂块中。 值得一提的是,人工合成的生长素类似物,也存在极性运输的特性。活性越强,极性运输越强[3]。 植物体内的IAA除了自由的活性状态外,还存在与糖、氨基酸结合的束缚状态(又称之为钝化)。如吲哚乙酸与葡萄糖结合为吲哚乙酸葡萄糖甙(葡萄糖甙),与蛋白质结合为吲哚乙酸-蛋白质复合物等,这类生长素常可占植物体中吲哚乙酸总量的50% 90%, 檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼檼 它们可能是植物解除过量吲哚乙酸毒性或避免 沿用至今。 4显微镜制造的发展 1725年,柯贝别尔氏所制造的显微镜才把灯光换成了反光镜,凿洞的桌子改成了带洞的载物台。从此显微镜不论在外形上还是在性能上都提高了一大步。 1744年,卡尔佩珀设计了第一台三只脚的台座式显微镜,它可以看作是现代显微镜的先驱。 1926年,布施设想出电子显微镜。1932年德国人鲁斯卡发明了世界上第一台电子显微镜,并因此获得了1953诺贝尔物理学奖。 1943年,德国人科诺尔和鲁斯卡首次对电子显微镜做出了重大的改革。后来很长一段时间内,电子显微镜的放大倍数一直没有增加,尽管如此,当时的电子显微镜已可观测到1/1000000mm的物体。 我国1965年试制成功第一台电子显微镜,放大倍数为20万倍,后来又制造出了80万倍的电子显微镜。 1982年,世界上第一台扫描隧道显微镜问世了。这是国际商业机器公司苏黎世实验室的宾尼格和罗尔及其同事们共同研制成功的世界上第一台新型表面分析仪器,使人类第一次能够观察到原子物质表面的排列状态,被国际公认为20世纪80年代世界十大科技之一。 为此,宾尼格和罗尔共同获得了1986年诺尔贝物理学奖。?
验证生长素的极性运输 一、教学设计思想 通过前面的学习,学生已知道植物生长素的发现过程,因而这次实验,是让学生体验发现生长素的过程和方法。本实验研究最好以小组为单位进行,学生可自愿结合组成若干小组,要求选出小组长负责本组的研究工作。 二、教学目标 1、知识目标:概述植物生长素的发现过程,理解生长素的极性运输方式。 2、能力目标: (1)学习用实验验证的方法来验证生长素的极性运输 (2)学会验证实验的一般方法和步骤。 3、情感态度和价值观目标: (1)培养学生科学探究能力,实验创新能力。 (2)通过实验过程中小组之间的合作,培养协作精神。 (3)体会科学理论往往在生产实践应用过程中需要探索解决的问题。 三、教学重点:概述植物生长素的发现过程,理解生长素的极性运输方式。 四、教学难点:掌握验证实验的一般方法和步骤。 五、学情分析 本实验是基于学生在学习了解植物生长素的发现过程的基础上进行实验的。学生往往比较有积极性,将课本上的科学知识融入具体的生活生产中这就是实验的出发点,即全面展示这节课的知识点,又要学生保持强烈的好奇心和求知欲,还原于生活,创设模拟科学家解决问题的方法。 六、课前准备: 加拿大飞蓬幼苗,琼脂块,生长素 七、教学过程: (一)、目的要求: 1、学会验证性实验的一般方法和步骤,培养科学探究能力,提高创新思维能力。 2、学会用验证的实验方法来验证生长素的极性运输。 3、理解生长素的极性运输,体会科学理论在应用到生产实践的过程中,往往也有许多要探索的问题。 (二)、实验原理:生长素(IAA)在胚芽鞘内只能从形态学上端(顶端)向下端(基端)运输,而不能倒转过来运输
收稿日期:2007-11-28 作者简介:常 莉(1981-),女,硕士研究生,专业方向:植物生理学及植物内源激素,E m ai:l s w c h angl@i 163 co m;通讯作者:薛建平,E m ai:l xu ej p2000@yahoo co m cn 。 生长素极性运输研究进展 常 莉,薛建平 (淮北煤炭师范学院生物系资源植物学安徽省重点实验室,淮北 235000) 摘 要:生长素极性运输与植物生长发育密切相关并受许多因素调控,生长素极性运输机理方面已取得较大进展,但仍有一些亟待解决的问题。研究植物生长素极性运输的生理机制及其调控具有十分重要的意义。通过了解生长素在植物生长发育中的作用,进而阐述生长素极性运输机理方面的研究进展。关键词:生长素;极性运输;植物生长发育中图分类号:Q 946 8851+ 1 文献标识码:A 文章编号:1008-9632(2008)06-0009-05 高等植物的生长发育受到植物激素的调控,目前公认的植物激素有五类,即生长素类、赤霉素类、细胞 分裂素类、乙烯和脱落酸。生长素早在1880年就被发现,也是最早被发现的一种植物激素。在植物体内普遍存在的生长素是吲哚乙酸(I AA ),天然生长素(如I AA )和合成生长素(如2,4 D 和NAA 等)在植物体中都表现出极性运输。目前已知的植物激素中只有生长素具有极性运输这一特征,因此很自然地就把植物的极性发育、分化、生长等生理现象与生长素的极性运输联系在一起。早期对生长素极性运输的研究主要采取施加极性运输抑制剂如萘基邻氨甲酰苯甲酸(NP A )、9 羟基 芴 9 羧酸(HFCA )、三碘苯甲酸(T I BA )和反式肉桂酸(CA )的方法。后来通过对生长素极性运输突变体的研究,从分子水平上为生长素极性运输的化学渗透偶联学说提供了新的证据,进一步发现了极性运输与植物的生长发育现象之间的联系。1 生长素极性运输的生理特点 生长素的极性运输是指生长素在植物体内由形态学的上端向形态学的下端单向运输的现象。在高等植物茎中,生长素由茎尖合成位点极性运输到茎基部的作用位点;在根的不同组织中则存在两种截然不同的运输方式:在中柱细胞中由根基向根尖的向顶式运输,表皮细胞中由根尖向根基的向基式运输。研究表明,生长素极性运输具有依赖于能量、需要O 2、对温度敏感、随年龄增加而减弱等生理特点,显然是一种需要消耗代谢提供能量的主动运输。其运输的速度比维管系统中的运输速度慢得多,又比物理扩散快约10倍,一般为5~20mm /h 。极性运输需要消耗能量,可逆浓度 梯度运输。一些化合物如2,3,5 三碘苯甲酸(T I BA )和奈基邻氨甲酞苯甲酸(NPA )等能抑制生长素极性运 输,缺氧会严重地阻碍生长素的运输。此外,生长素又有自动抑制现象,即先发育的器官通过其合成并向外输出的生长素抑制后发育期器官生长素的输出。2 生长素极性运输与植物的生长和发育 一般认为生长素主要在植物叶原基、幼叶以及发育的种子等部位合成。在这些器官中合成的生长素需要通过极性运输到达靶细胞才能调节植物的生长和发育,因此生长素极性运输广泛参与植物的叶片发育、花的分化、维管的分化、胚胎发育、光形态建成以及侧根的发育等。 2 1 生长素极性运输和叶片发育 叶片是植物进行光合作用和呼吸作用的主要器官,对植物生命活动起着重要作用。形态学上,成熟叶片具有三个轴:近远轴,腹背轴和中心 两侧轴。体内的遗传机制和体外的环境因子正是沿着这些轴向来 调控叶片发育和形态建成[1] 。早期叶片发育可被人为的分为三个阶段:叶原基的起始,腹背性的建立和叶片的延展 [1] 。在这些阶段中发生的任何突变都有可能 产生叶片发育和叶片形态的缺陷。在研究离体芥菜胚和小麦胚的培养中,生长素极性运输抑制剂导致了芥菜胚筒状子叶和小麦胚柄与芽分生组织之间的伸长。L i u [1] 等提出:生长素极性运输可以使将来发育成子叶的两个区域中生长素达到合适的浓度,从而有利于子叶原基的发端而形成两侧对称的子叶。后来在研究离体培养的二月兰等的叶或花柄外植体而进行的不定芽分化,加入生长素极性运输抑制剂后,即有喇叭状叶和
必修3 第3章植物的激素调节(人教新课标) 第1节植物生长素的发现 【思维导图】 【微试题】 1 如图所示,①②③为对胚芽鞘作不同处理的实验,④为一植株被纸盒罩住,纸盒的一侧开口,有单侧光照。下列对实验结果的描述,正确的是( ) A.①②中胚芽鞘向光弯曲生长,③中胚芽鞘背光弯曲生长 B.①中胚芽鞘直立生长,②③中胚芽鞘向光弯曲生长 C.④中如果固定植株,旋转纸盒,一段时间后,植株向左弯曲生长 D.④中如果将纸盒和植株一起旋转,则植株弯向盒开口方向生长 【方法规律】用“两看法”判断植物生长弯曲状况 (1)一看胚芽鞘尖端以下的部分能否获得生长素:①能则生长;②不能则既不生长也不弯曲。 (2)二看胚芽鞘尖端以下部分生长素的分布是否均匀:①若均匀则直立生长;②若不均匀则弯
曲生长。 2 取生长状态一致的燕麦胚芽鞘,分为a、b、c、d四组。将a、b两组胚芽鞘尖端下方的一段切除,再从c、d两组胚芽鞘中的相应位置分别切取等长的一段,并按下图中所示分别接入a、b两组胚芽鞘被切除的位置,得到a′、b′两组胚芽鞘。然后用单侧光照射,发现a′组胚芽鞘向光弯曲生长,b′组胚芽鞘无弯曲生长,其原因是( ) A.c组尖端能合成生长素,d组尖端不能 B.a′组尖端能合成生长素,b′组尖端不能 C.c组尖端的生长素能向胚芽鞘基部运输,d组尖端的生长素不能 D.a′组尖端的生长素能向胚芽鞘基部运输,b′组尖端的生长素不能 【微探究】 (1)若随太空飞船进入太空中,植物还能发生极性运输吗?还具有向光性吗? 提示:极性运输由植物的遗传特性决定,即使在太空中也会发生极性运输,也具有向光性。(2)若加入呼吸抑制剂,则植物生长素的运输会受到影响,由此说明生长素的运输方式是什么? 提示:主动运输。 3 [2014全国高考] 为了验证单侧光照射会导致燕麦胚芽鞘中生长素分布不均匀这一结论,需要先利用琼脂块收集生长素,之后再测定其含量。假定在单侧光照射下生长素的不均匀分布只与运输有关,下列收集生长素的方法(如图示)中,正确的是( )
生长素在根部的几种 运输方式
生长素在根部运输的几种方式 茎中生长素的运输方向是形态学上端到下端的极性运输,那么,在植物根中生长素的运输方向是什么呢? 按照目前流行的各种高中生物教辅资料中,在解释把植物横放后根的向重力性时生长原因时,一般都认为:是根尖产生了生长素,在重力的作用下,由根尖的远地侧向近地侧横向运输,然后再向根基部方向运输,由于根对生长素很敏感,故抑制了近地侧根细胞的生长,使远地侧根细胞生长速度较快而出现向重力性生长。也就是说,根中生长素是由根尖向根基部的极性运输。然而,这种说法是不正确的。 1.根中生长素的来源 IAA主要是在植物的顶端分生组织中合成的,合成后即被运输到植株各部和根中。IAA的运输是单方向的,只能从植物顶端往下运输而不能反过来从下面往顶端运输,所以称为极性运输。根据根向重力性Evans-Moore模型,根直立生长时茎尖运向根冠的IAA在根中四周平均分布。高中教材上也提到,生长素主要产生于幼嫩的芽、叶,发育中的种子等。生长素在植物中分布很广,但以生长旺盛的器官和组织如正在生长的茎尖和根尖的分生组织、胚芽鞘、受精后的子房和幼嫩的种子等含量较多,而在衰老的组织和器官中含量甚少。从这里可以这样理解:根尖尤其是根尖分生组织中的较多生长素不是根尖产生的,而是茎尖产生后向下运输一直运输到根尖,并作用于根尖的。 2.根中生长素的极性运输 植物根中的生长素也表现极性运输的性质,不过是由根基部向根尖方向的运输,即向顶运输。 目前,对根的向重性普遍是这样解释的,也就是Evans(1996)提出的根向重力性Evans-Moore模型:IAA在地上部分合成,经维管系统运输到根,当根尖处于与重力线方向平行时,根冠细胞中淀粉体沉降在柱细胞的底端,此时Ca2+和运到根冠的IAA向四周平均分配。然后IAA再经根皮层向基方向运至根伸长区,以促进伸长区细胞均衡伸长,使根仍与重力线方向平行生长(图A)。但当根处于水平方向时(图B),淀粉体沉降至柱细胞下侧,从而促进Ca2+与IAA在下侧释放。Ca2+还增强IAA进入向基性的运输流,使IAA更多地经皮层运输到根的下侧,并在下侧积累。这种超最适浓度的IAA 会抑制根下侧的伸长,从而引起根向下弯曲的运动反应。
专题五生命系统的稳态及调节 整合考点13 “生长靠我”的生长素
1.理清生长素的“5”个易错点 (1)生长素的产生部位在尖端,其合成不需要光。 (2)感光部位在尖端,单侧光照射尖端会引起生长素分布不均匀。 (3)横向运输是在尖端完成的,但发生作用的部位在尖端下面的一段。 (4)生长素不能透过云母片,但能透过琼脂。 (5)若无尖端,外源生长素不对称放置,也会引起生长素分布不均匀 2.理解生长素的三类运输方式 (1)极性运输 ????? 方向:形态学上端―→形态学下端方式:主动运输 (2)非极性运输:在成熟组织中,生长素通过韧皮部的筛管进行非极性运输。 (3)横向运输:在某些刺激(如单侧光、地心引力等)影响下,生长素在能感受刺激的部位(如胚芽鞘的尖端)发生横向运输(不接受刺激的部位不能发生横向运输)。 3.辨明生长素作用两重性的含义 (1)高浓度与低浓度:当生长素浓度小于i 时均为“低浓度”,高于i 时才会抑制植物生长,称为“高浓度”。 (2)OH 段表明:随生长素浓度升高,促进生长作用增强。 (3)HC 段表明:随生长素浓度升高,促进生长作用减弱(但仍为促进生长)。 (4)最适浓度:H 点表示促进生长的最适浓度为g 。 (5)“阈值”:C 点表示促进生长的“阈值”,即大于C 处生长素浓度,将会使生长受抑制。 生长素的发现、产生、分布和运输 1.(2016·全国卷Ⅲ)为了探究生长素的作用,将去尖端的玉米胚芽鞘切段随机分成两组,实验组胚芽鞘上端一侧放置含有适宜浓度IAA 的琼脂块,对照组胚芽鞘上端同侧放置不含IAA 的琼脂块,两组胚芽鞘下段的琼脂块均不含IAA 。两组胚芽鞘在同样条件下,在黑暗中放置一段时间后,对照组胚芽鞘无弯曲生长,实验组胚芽鞘发生弯曲生长,如图所述。根
龙源期刊网 https://www.doczj.com/doc/f110412426.html, 浅议植物生理学中生长素在根中的运输问题作者:李积胜 来源:《科技资讯》2014年第34期 摘要:植物重要激素生长素与植物正在发育密切相关。有关生长素在植物根中运输的相 关信息,在植物生理学教材植物生长物质这一章节中并不完整。长期以来学生对生长素运输的认识不深入,导致没有把生长素运输及生长素分布与生长素的生理功能相结合,使得学生在学习中对生长素生理作用产生了疑惑。在植物生理教材中添加这样的内容有助于学生更好的理解这样生理过程,所以完整的补充这一知识点有利于学生全面的认识这一植物激素。 关键词:植物生理学植物激素生长素运输模式 中图分类号:Q946.8 文献标识码:A 文章编号:1672-3791(2014)12(a)-0157-01 在当代本科生教育中,《植物生理学》一直是生物类学生非常重要的一门专业课,也是生物类考研考生大多会遇到的一门课程。多年来植物生理学教科书的不同版本相继推出,这些教材无论从深度和广度都不断提高。但是涉及到植物生长物质这一章节中植物重要激素生长素,很少提及生长素在植物根中运输模式是怎样的。尽管国外植物生理学教科书已经及时更新了植物根中生长素运输的相关内容,但还缺少一个完整的生长素运输模型,这不利于学生全面理解和掌握植物重要激素生长素在细胞内的调节机制和作用模式。完整的补充这一知识点有利于学生全面的认识这一植物激素,了解生长素运输对生长素发挥作用所带来的影响。因此,建议及时补充植物生理学中生长素在植物根中的运输模式。 1 生长素运输对于植物生长的重要意义 生长素主要在植物叶原基、嫩叶和茎尖等生命力旺盛的部位合成,随后以一种特殊的极性转运方式(自植物形态学上端向形态学下端运输)将其运输到目的地发挥生物学功能。早期在植物中使用生长素运输抑制剂发现,生长素运输在调节植物器官发育、根和茎分支、植物可逆的向光性和向地性等方面的作用尤为突出。生长素通过极性运输维持其在植物体内的梯度性分布,是植物形态建成的主要决策者[1]。 2 生长素在植物根中运输模式 极性运输是生长素的一个重要特征。传统意义上的生长素极性运输是指生长素从植株的茎尖生长点和幼叶等组织向下部组织的单向运输。近十几年来,对生长素的极性运输有了更深入的研究,已知植物中不仅存在着从茎尖向根的极性运输,还具有在组织局部的极性运输。例如,在根尖存在着生长素的环流,从地上部分通过维管组织运输来的生长素经过根尖中柱细胞间的单向运输,向根尖静止中心聚集,然后又通过表皮细胞和侧根冠细胞向根尖的伸长区运
有关生长素发现史知识点考题例析 生长素的发现是经典生物学实验之一,它遵循了科学研究的基本思路。通过分析有关生长素的发现实验史,可以掌握其中的科学方法和重要实验结论。本文拟从三个方面,对发现史中的相关知识点进行了剖析,以期对学生探究意识和实践能力的培养有所帮助。 一、与“几个重要结论”有关的实验分析 1.要点把握: ⑴产生生长素及感受单侧光刺激的部位:胚芽鞘尖端(仅该部位能感受这一外界刺激,而其他部位对单侧光的刺激无反应)。 ⑵生长和向光弯曲的部位:胚芽鞘尖端下面的一段。 ⑶单侧光只影响生长素的分布,而光照不是生长素合成的必要条件。 2.知识延伸: 胚芽鞘向光弯曲的五要点:⑴生长素在尖端有横向运输的能力;⑵生长素在尖端有极性运输的能力; ⑶生长素作用机理是促进细胞的纵向伸长。⑷向光性的内因是生长素的不均匀分布。⑸向光性的外因:单侧光的影响,胚芽鞘的生长素背光侧多→生长快,而向光侧少→生长慢。 例1.科学家研究胚芽鞘向光弯曲现象,逐渐揭示了发生这种应激反应的一系列因果相关事件。下列各项中,按因果相关事件顺序排列的是() a.胚芽鞘尖端合成生长素 b.胚芽鞘尖端感受光刺激c.胚芽鞘向光弯曲生长 d.生长素在背光一侧分布较多e.背光一侧细胞生长较快 f.单侧光照射胚芽鞘尖端 【解析】本题考查了学生对生长素产生部位、感受光刺激的部位、引起植物向光性现象的原因等知识的理解和应用。胚芽鞘向光弯曲的前提,即胚芽鞘尖端可以合成生长素。在单侧光的照射下,尖端感受光的刺激,导致生长素运输不均。使背光侧生长素的含量多于向光侧,背光侧细胞生长得快,向光侧细胞生长的较慢,最后表现为向光弯曲生长 【答案】D 二、与“几种常用材料”有关的实验分析