一、序列
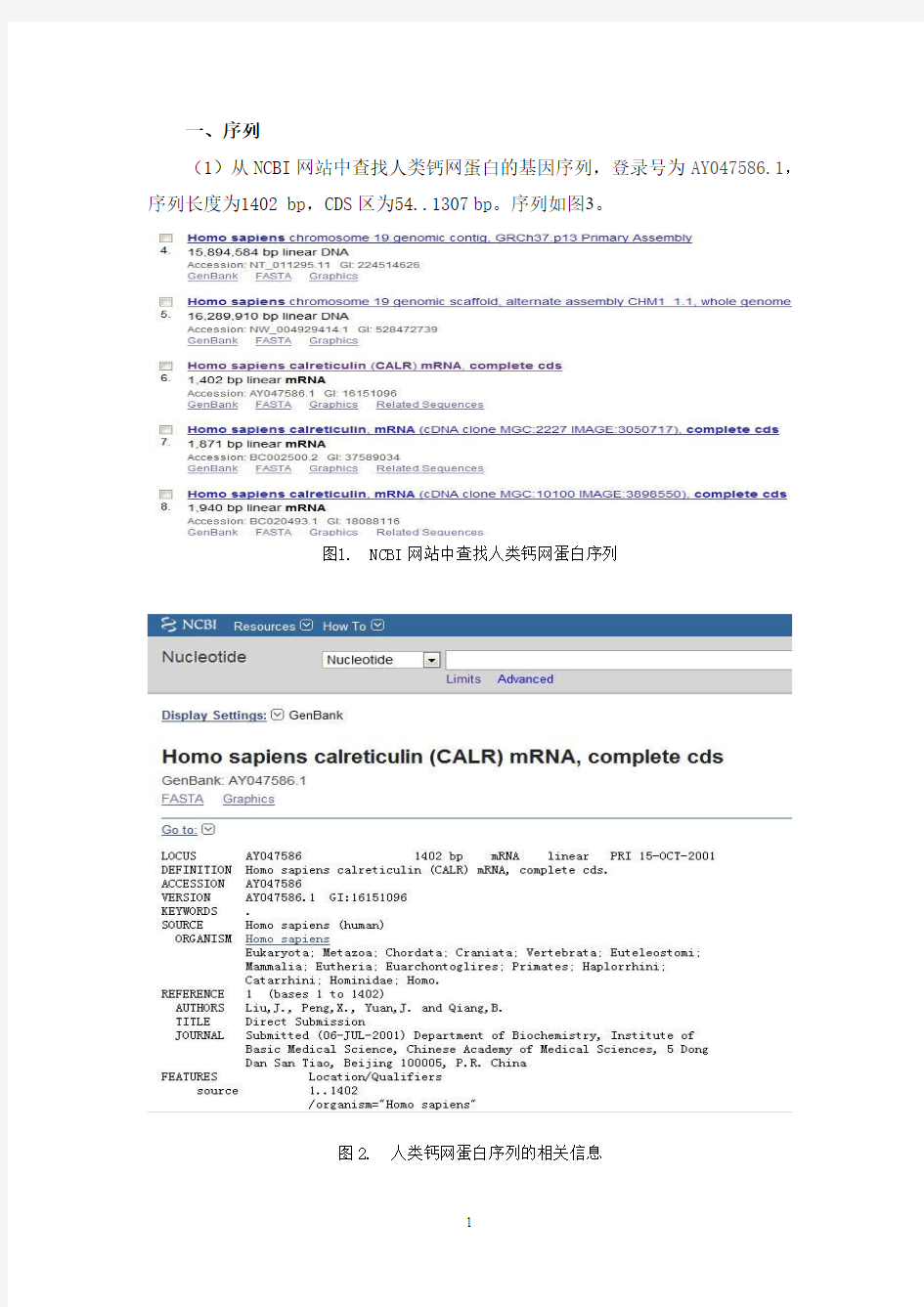
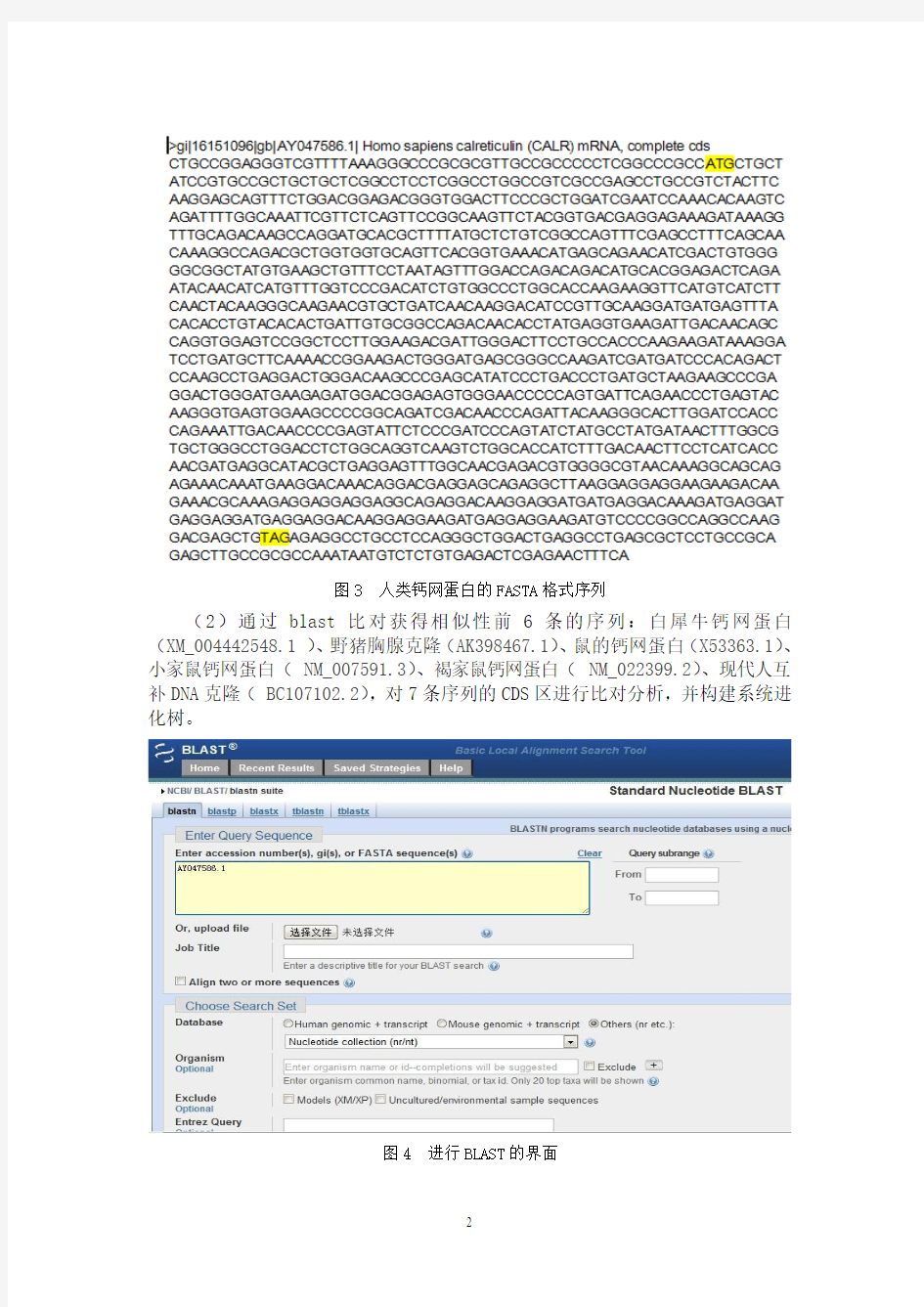
(1)从NCBI网站中查找人类钙网蛋白的基因序列,登录号为AY047586.1,序列长度为1402 bp,CDS区为54..1307bp。序列如图3。
图1. NCBI网站中查找人类钙网蛋白序列
图2. 人类钙网蛋白序列的相关信息
图3 人类钙网蛋白的FASTA格式序列
(2)通过blast比对获得相似性前6条的序列:白犀牛钙网蛋白(XM_004442548.1 )、野猪胸腺克隆(AK398467.1)、鼠的钙网蛋白(X53363.1)、小家鼠钙网蛋白( NM_007591.3)、褐家鼠钙网蛋白( NM_022399.2)、现代人互补DNA克隆( BC107102.2),对7条序列的CDS区进行比对分析,并构建系统进化树。
图4 进行BLAST的界面
图5 BLAST之后的结果
图6 BLAST之后的结果
图7 MAGA的运行结果
图8 MEGA的运行结果
图9 系统进化树
二、对人类钙网蛋白的蛋白质进行一级结构的预测
从NCBI中搜索人类钙网蛋白的蛋白序列,其登录号为AAL13126.1,序列如图所示:
图10 人类钙网蛋白的蛋白序列
通过protparam(https://www.doczj.com/doc/5715348065.html,/tools/protparam.html)对人类钙网蛋白的蛋白质的基本理化性质进行预测,结果显示该蛋白编码氨基酸数目为417,相对分子质量为48141.5 Da,理论pI值4.29。
图11 protparam的首页
图12 蛋白质的氨基酸数目.相对分子质量.理论pI值.氨基酸组成
图13 蛋白质的正/负电荷残基数.分子式.总原子数
图14 蛋白质的消光系数.半衰期.不稳定系数.脂肪系数.总平均亲水性
三、对该蛋白质二级结构进行预测(亲疏水性、跨膜区、结构域等)
(1)通过protscale (https://www.doczj.com/doc/5715348065.html,/tools/protscale.html)网站进行亲疏水性预测。
图15 亲疏水性工具protscale首页
图16 亲疏水性预测的结果
图17 亲疏水性预测的结果
(2)通过http://www.cbs.dtu.dk/services/TMHMM-2.0这个网站,对该蛋白质进行跨膜区预测
图19 跨膜区工具TMHMM首页
图19 跨膜区预测结果
(3)通过(http://smart.embl-heidelberg.de/)这个网站,对该蛋白质进行结构域预测
图19 结构域工具smart页面
图20 结构域预测结果
四、通过(https://www.doczj.com/doc/5715348065.html,/)这个网站,对该蛋白质三级结构进行预测
图21 三维工具swiss-modle页面
图22 三级结构预测结果
个人收获
生物信息学通俗的说法就是利用数学和计算机知识来处理生物数据,在这一个学期内的学习中,不仅学习到了有关生物信息学的一些理论知识,而且也使我们接触到了怎样用现代技术来处理得到的数据,每一步做出来的东西都让我们很惊讶也很惊喜,原来生物世界是这么美妙。同时,通过它也使人类对生物的研究更提升了一个水平。在以后的学习生活中,我会一如既往的保持原有的严谨性,不仅学习好我的理论知识,还锻炼好我的动手能力。
常用数据库 在DNA序列方面有GenBank、EMBL和等 在蛋白质一级结构方面有SWISS-PROT、PIR和MIPS等 在蛋白质和其它生物大分子的结构方面有PDB等 在蛋白质结构分类方面有SCOP和CATH等 生物信息学的主要研究内容 1、序列比对(Alignment) 基本问题是比较两个或两个以上符号序列的相似性或不相似性。序列比对是生物信息学的基础,非常重要。两个序列的比对有较成熟的动态规划算法,以及在此基础上编写的比对软件包BLAST和FASTA,可以免费下载使用。这些软件在数据库查询和搜索中有重要的应用。 2、结构比对 基本问题是比较两个或两个以上蛋白质分子空间结构的相似性或不相似性。已有一些算法。 3、蛋白质结构预测,包括2级和3级结构预测,是最重要的课题之一 从方法上来看有演绎法和归纳法两种途径。前者主要是从一些基本原理或假设出发来预测和研究蛋白质的结构和折叠过程。分子力学和分子动力学属这一范畴。后者主要是从观察和总结已知结构的蛋白质结构规律出发来预测未知蛋白质的结构。同源模建(Homology)和指认(Threading)方法属于这一范畴。虽然经过30余年的努力,蛋白结构预测研究现状远远不能满足实际需要。 4、计算机辅助基因识别(仅指蛋白质编码基因)。最重要的课题之一 基本问题是给定基因组序列后,正确识别基因的范围和在基因组序列中的精确位置.这是最重要的课题之一,而且越来越重要。经过20余年的努力,提出了数十种算法,有十种左右重要的算法和相应软件上网提供免费服务。原核生物计算机辅助基因识别相对容易些,结果好一些。从具有较多内含子的真核生物基因组序列中正确识别出起始密码子、剪切位点和终止密码子,是个相当困难的问题,研究现状不能令人满意,仍有大量的工作要做。 5、非编码区分析和DNA语言研究,是最重要的课题之一 在人类基因组中,编码部分进展总序列的3~5%,其它通常称为“垃圾”DNA,其实一点也不是垃圾,只是我们暂时还不知道其重要的功能。分析非编码区DNA 序列需要大胆的想象和崭新的研究思路和方法。DNA序列作为一种遗传语言,不仅体现在编码序列之中,而且隐含在非编码序列之中。 6、分子进化和比较基因组学,是最重要的课题之一 早期的工作主要是利用不同物种中同一种基因序列的异同来研究生物的进化,构建进化树。既可以用DNA序列也可以用其编码的氨基酸序列来做,甚至于可通过相关蛋白质的结构比对来研究分子进化。以上研究已经积累了大量的工作。近年来由于较多模式生物基因组测序任务的完成,为从整个基因组的角度来研究分子进化提供了条件。 7、序列重叠群(Contigs)装配 一般来说,根据现行的测序技术,每次反应只能测出500或更多一些碱基对的序列,这就有一个把大量的较短的序列全体构成了重叠群(Contigs)。逐步把它们拼接起来形成序列更长的重叠群,直至得到完整序列的过程称为重叠群装配。拼接EST数据以发现全长新基因也有类似的问题。已经证明,这是一个NP-完备
生物信息学试题 1、构建分子系统树得主要方法有哪些?并简要说明构建分子进化树 得一般步骤。(20分) 答:(1)构建进化树得方法包括两种:一类就是序列类似性比较,主要就是基于氨基酸相对突变率矩阵(常用PAM250)计算不同序列差异性积分作为它们得差异性量度(序列进化树);另一类在难以通过序列比较构建序列进化树得情况下,通过蛋白质结构比较包括刚体结构叠合与多结构特征比较等方法建立结构进化树 (2)序列比对——选取所需序列——软件绘制 具体如下: a测序获取序列或者在NCBI上搜索所需得目得序列 b在NCBI上做blast:比对相似度较高得基因,并以fast格式下载,整合在*txt文档中。 c比对序列,比对序列转化成*meg格式 d打开保存得*meg格式文件,构建系统进化树 2、氨基酸序列打分矩阵PAM与BLOSUM中序号有什么意义?它们各自 得规律就是什么?(10分) (1)PAM矩阵:基于进化得点突变模型,如果两种氨基酸替换频繁,说明自然界接受这种替换,那么这对氨基酸替换得分就高。一个PAM就就是一个进化得变异单位, 即1%得氨基酸改变。 BLOSUM矩阵:首先寻找氨基酸模式,即有意义得一段氨基酸片断,分别比较相同得氨基酸模式之间氨基酸得保守性(某种氨基酸对另一种氨基酸得取代数据),然后,以所有60%保守性得氨基酸模式之间得比较数据为根据,产生BLOSUM60;以所有80%保守性得氨基酸模式之间得比较数据为根据,产生BLOSUM80。
(2)PAM用于家族内成员相比,然后把所有家族中对某种氨基酸得比较结果加与在一起,产生“取代”数据(PAM-1 );PAM-1自乘n次,得PAM-n。 PAM-n中,n 越小,表示氨基酸变异得可能性越小;相似得序列之间比较应该选用n值小得矩阵,不太相似得序列之间比较应该选用n值大得矩阵。PAM-250用于约 20%相同序列之间得比较。 BLOSUM-n中,n越小,表示氨基酸相似得可能性越小;相似得序列之间比较应该选用 n 值大得矩阵,不太相似得序列之间比较应该选用n值小得矩阵。BLOSUM-62用来比较62%相似度得序列,BLOSUM-80用来比较80%左右得序列。 3、蛋白质三维结构预测得主要方法有哪些?试选择其中得一种方 法,说明蛋白质三维结构预测得一般步骤。(10分) (1) a同源建模(序列相似性低于30%得蛋白质难以得到理想得结构模型 b折叠识别(已知结模板得序列一致率小于25%) c从头预测得方法(无已知结构蛋白质模板)。 (2) 4、您所熟悉得生物信息学软件有哪些?请选择其中得至少一种软 件,结合自己得研究课题,谈谈您所选择软件得基本原理,使用
基因组测序基础知识 ㈠De Novo测序也叫从头测序,是首次对一个物种的基因组进行测序,用生物信息学的分析方法对测序所得序列进行组装,从而获得该物种的基因组序列图谱。 目前国际上通用的基因组De Novo测序方法有三种: 1. 用Illumina Solexa GA IIx 测序仪直接测序; 2. 用Roche GS FLX Titanium直接完成全基因组测序; 3. 用ABI 3730 或Roche GS FLX Titanium测序,搭建骨架,再用Illumina Solexa GA IIx 进行深度测序,完成基因组拼接。 采用De Novo测序有助于研究者了解未知物种的个体全基因组序列、鉴定新基因组中全部的结构和功能元件,并且将这些信息在基因组水平上进行集成和展示、可以预测新的功能基因及进行比较基因组学研究,为后续的相关研究奠定基础。 实验流程: 公司服务内容 1.基本服务:DNA样品检测;测序文库构建;高通量测序;数据基本分析(Base calling,去接头, 去污染);序列组装达到精细图标准 2.定制服务:基因组注释及功能注释;比较基因组及分子进化分析,数据库搭建;基因组信息展 示平台搭建 1.基因组De Novo测序对DNA样品有什么要求?
(1) 对于细菌真菌,样品来源一定要单一菌落无污染,否则会严重影响测序结果的质量。基因组完整无降解(23 kb以上), OD值在1.8~2.0 之间;样品浓度大于30 ng/μl;每次样品制备需要10 μg样品,如果需要多次制备样品,则需要样品总量=制备样品次数*10 μg。 (2) 对于植物,样品来源要求是黑暗无菌条件下培养的黄化苗或组培样品,最好为纯合或单倍体。基因组完整无降解(23 kb以上),OD值在1.8~2.0 之间;样品浓度大于30 ng/μl;样品总量不小于500 μg,详细要求参见项目合同附件。 (3) 对于动物,样品来源应选用肌肉,血等脂肪含量少的部位,同一个体取样,最好为纯合。基因组完整无降解(23 kb以上),OD值在1.8~2.0 之间;样品浓度大于30 ng/μl;样品总量不小于500 μg,详细要求参见项目合同附件。 (4) 基因组De Novo组装完毕后需要构建BAC或Fosmid文库进行测序验证,用于BAC 或Fosmid文库构建的样品需要保证跟De Novo测序样本同一来源。 2. De Novo有几种测序方式 目前3种测序技术 Roche 454,Solexa和ABI SOLID均有单端测序和双端测序两种方式。在基因组De Novo测序过程中,Roche 454的单端测序读长可以达到400 bp,经常用于基因组骨架的组装,而Solexa和ABI SOLID双端测序可以用于组装scaffolds和填补gap。下面以solexa 为例,对单端测序(Single-read)和双端测序(Paired-end和Mate-pair)进行介绍。Single-read、Paired-end和Mate-pair主要区别在测序文库的构建方法上。 单端测序(Single-read)首先将DNA样本进行片段化处理形成200-500bp的片段,引物序列连接到DNA片段的一端,然后末端加上接头,将片段固定在flow cell上生成DNA簇,上机测序单端读取序列(图1)。 Paired-end方法是指在构建待测DNA文库时在两端的接头上都加上测序引物结合位点,在第一轮测序完成后,去除第一轮测序的模板链,用对读测序模块(Paired-End Module)引导互补链在原位置再生和扩增,以达到第二轮测序所用的模板量,进行第二轮互补链的合成测序(图2)。 图1 Single-read文库构建方法图2 Paired-end文库构建方法
上海师范大学实验报告 实验二 一、实验原理 答:利用Blast全球联网数据库,对输入的序列进行生物信息学分析,给出与输入序列相关性最大的对应的基因信息,比较两者的同源性。 二、操作步骤 答:(1)先打开网址https://www.doczj.com/doc/5715348065.html,/ (2)点击右边的Blast链接,打开Blast数据库,进入Blast界面 (3)在Basic Blast中选择nucleotide blast (4)在对话框中输入核苷酸序列,在choose search set下的Database选项中选择Others (nr etc.) (5)把网页拉到最下方,点击Blast按钮 (6)在Descriptions 栏下找到Max ident 百分率最高的序列名称 (7)再往下拉,找到Alignments项下第一个序列,可以找到输入序列相关信息 (8)点击Accession,即能找到更多输入序列的相关信息。 1. tttcactcca tagttactcc ccaggtga 1.1它属于哪类生物? 答:属于Hepatitis C virus (丙型肝炎病毒) 1.2它属于哪类基因? 答:属于non-structural protein 5B gene 1.3它在该基因的什么位置? 答:它在该基因的第749-776这个位置。 1.4它与你搜索到的序列的同源性(Identities)是多少? 答:同源性100% 2.(1)ccacccactg aaactgcaca gacaaatttg tacataagag 1.1它属于哪类生物? 答:属于Influenza A virus (A/chicken/Iran261/01(H9N2)) hemagglutinin (HA) gene (A型流感病毒,A型伊朗型261鸡流感病毒,H9N2病毒,血细胞凝集素抗原基因为依据) 1.2它属于哪类基因? 答:属于ssRNA negative-strand viruses Orthomyxoviridae (单链RNA,负义链病毒,正粘病毒科) 1.3它在该基因的什么位置? 答:它在该基因的第1-40这个位置 1.4它与你搜索到的序列的同源性(Identities)是多少?
水稻瘤矮病毒(RGDV)外层衣壳蛋白 P8的同源模建 高芳銮(Raindy) 同源模建(homology modeling) ,也叫比较模建(Compatative modeling),其前提是一个或多个同源蛋白质的结构已知,当两个蛋白质的序列同源性高于35%,一般情况下认为它们的三维结构基本相同;序列同源性低于30%的蛋白质难以得到理想的结构模型。同源模建是目前最为成功且实用的蛋白质结构预测方法, SWISS-MODEL 是由SwissProt 提供的目前最著名的蛋白质三级结构预测服务器,创建于1993年,面向全世界的生物化学与分子生物学研究工作者提供免费的自动模建服务。SWISS-MODEL 服务器提供的同源模建有两种工作模式:首选模式(First Approach mode)和 项目模式(Project mode)。 本实例以RGDV P8蛋白为研究对象采用首选模式进行同源模建。 图1 SWISS-MODEL 的主界面 操作流程如下: 1.选择模式 单击左侧的“MENU ”菜单下方的“First Approach mode ”,右侧窗口自动SWISS-MODEL 工作窗口,在相应文本框中分别输入的E-mail 、项目标题、待模建的蛋白质序列,SWISS-MODEL 支持以FASTA 格式直接输入或提交UniProt 的登录号,如图2所示。 《生物信息学分析实践》样 稿
图2 SWISS-MODEL 的序列提交页面 2.参数设置 当前版本只有一个选项可设置,如果用户需要使用指定的模板,可在“Use a specific template ”后的输入框填入ExPDB 晶体图像数据库中的模板代码,其格式为“PDBCODE+ChainID ”,如“1uf2P ”。本例不使用指定模板,默认留空。完毕,点击“Submit Modeling Request ”提交模建请求,服务器返回提交成功的提示,如图3所示: 图3 成功提交 SWISS-MODEL WORKSPACEW 页面会自动刷新,直至模建完成,如图4所示,同时模建结果也会发送到指定的邮箱。 3结果解读 点击下图右上方的“Print/Save this page as ”后的图标,可以将整个结果以PDF 文档格式保存到本地计算机中。模建结果给出了五个部分的信息:模建详情(Model Details)、比对信息(Alignment)、模建评价 (Anolea/Gromos/Verify3D)、模建日志(Modelling log)、模板选择日志(Template Selection Log)。 《生物信息学分析实践》样稿
(生物科技行业)功能基因的克隆及生物信息学分析
功能基因的克隆及其生物信息学分析 摘要:随着多种生物全基因组序列的获得,基因组研究正从结构基因组学(structuralgenomics)转向功能基因组学(functionalgenomics)的整体研究。功能基因组学利用结构基因组学研究获得的大量数据与信息评价基因功能(包括生化功能、细胞功能、发育功能、适应功能等),其主要手段结合了高通量的大规模的实验方法、统计和计算机分析技术[1],它代表了基因分析的新阶段,已成为21世纪国际生命科学研究的前沿。功能基因组学是利用基因组测序获得的信息和产物,发展和应用新的实验手段,通过在基因组或系统水平上全面分析基因的功能,使生物学研究从对单一基因或蛋白的研究转向多个基因或蛋白同时进行系统的研究,是在基因组静态的组成序列基础上转入对基因组动态的生物学功能学研究[2]。如何研究功能基因,也成为我们面临的一个课题,本文就克隆和生物信息学分析在研究功能基因方面的应用做一个简要的阐述。 关键词:功能基因、克隆、生物信息学分析。 1.功能基因的克隆 1.1图位克隆方法 图位克隆又称定位克隆,它是根据目标基因在染色体上确切位置,寻找与其紧密连锁的分子标记,筛选BCA克隆,通过染色体步移法逐步逼近目的基因区域,根据测序结果或用BAC、YAC克隆筛选cDNA表达文库寻找候选基因,得到候选基因后再确定目标基因。优点是无需掌握基因产物的任何信息,从突变体开始,逐步找到基因,最后证实该基因就是造成突变的原因。通过图位克隆许多
控制质量性状的单基因得以克隆,最近也有报道某些控制数量性状的主效基因(控制蕃茄果实大小的基因克隆[3]、控制水稻成熟后稻谷脱落基因克隆[4]以及小麦VRN2基因克隆[5]等)也通过图位克隆法获得。 1.2同源序列克隆目的基因 首先根据已知的基因序列设计PCR引物,在已知材料中扩增到该片段,并经克隆测序验证,利用放射性同位素标记或其他非同位素标记该PCR片段作为探针,与待研究材料的cDNA文库杂交,就可以获得该基因cDNA克隆,利用克隆进一步筛选基因组文库,挑选阳性克隆,亚克隆并测序,从中就可以筛选到该基因的完整序列。 1.3结合连锁和连锁不平衡的分析方法 结合连锁和连锁不平衡的分析方法是未知基因克隆研究领域发展的新方向[6]。(Linkagedisequilibrium,LD)。与连锁分析不同,连锁不平衡分析可以利用自然群体中历史发生的重组事件。历史上发生的重组使连锁的标记渐渐分布到不同的同源染色体上,这样就只有相隔很近的标记才能不被重组掉,从而形成大小不同的单倍型片段(Haplotypeblock)。这样经过很多世代的重组,只有相隔很近的基因,才能仍处在相同的原始单倍型片段上,基因间的连锁不平衡才能依然存在。所以基于连锁不平衡分析,可以实现目的基因的精细定位。林木大多为自由授粉的异交物种,所以连锁不平衡程度很低,林木基因组中的LD可能会仅局限于非常小的区域,这就为目的基因的精细定位提供了可能,结合SNP检测技术,科学家甚至可以将效应位点直接与单个的核苷酸突变关联起来,进行数量性状寡核苷酸
生物信息学实验指导 适用专业:生物技术与制药大类 生物技术 编写:解增言 生物信息学院 2014年9月
目录 实验1 在线BLAST同源序列查询 (3) 实验2 本地BLAST同源序列查询 (8) 实验3 利用ClustalX与MEGA进行多序列比对与分子系统发生树构建 (10) 实验4 利用RNAfold预测RNA二级结构 (14) 实验5 Pfam蛋白质结构域分析 (17) 实验6 利用PSSpred预测蛋白质二级结构 (19) 实验7 利用Cn3D和RasMol分析蛋白质三级结构 (21) 实验8 利用GO及EST数据分析基因功能 (24)
实验1 在线BLAST同源序列查询 一、实验目的 1.了解同源序列查询的原理和用途; 2.掌握利用NCBI在线BLAST工具查找同源序列的方法。 二、实验原理 在生物学种系发生理论中,若两个或多个结构具有相同的祖先,则称它们同源(homologous)。分子生物学中的同源指两条序列来自于一条共同的祖先序列。一般来说,相似超过一定程度的序列具有同源性。在生物信息学研究中,常用序列比对(alignment)来研究序列的同源性以及推测物种之间的关系。 最常见的比对是蛋白质序列之间或核酸序列之间的两两比对,通过比较两个序列之间的相似区域和保守性位点,寻找二者可能的分子进化关系。进一步的比对是将多个蛋白质或核酸同时进行比较,寻找这些有进化关系的序列之间共同的保守区域或位点,从而探索导致它们产生共同功能的序列模式。此外,还可以把蛋白质序列与核酸序列相比来探索核酸序列可能的表达框架;把蛋白质序列与具有三维结构信息的蛋白质相比,从而获得蛋白质折叠类型的信息。 比对还是数据库搜索算法的基础,将查询序列与整个数据库]的所有序列进行比对,从数据库中获得与其最相似序列的已有的数据,能最快速的获得有关查询序列的大量有价值的参考信息,对于进一步分析其结构和功能都会有很大的帮助。近年来随着生物信息学数据大量积累和生物学知识的整理,通过比对方法可以有效地分析和预测一些新发现基因的功能。 序列两两比对 序列比对的理论基础是进化学说,如果两个序列之间具有足够的相似性,就推测二者可能有共同的进化祖先,经过序列内残基的替换、残基或序列片段的缺失、以及序列重组等遗传变异过程分别演化而来。序列相似和序列同源是不同的概念,序列之间的相似程度是可以量化的参数,而序列是否同源需要有进化事实的验证。在残基-残基比对中,可以明显看到序列中某些氨基酸残基比其它位置上的残基更保守,这些信息揭示了这些保守位点上的残基对蛋白质的结构和功能是至关重要的,例如它们可能是酶的活性位点残基,形成二硫键的半胱氨酸残基,与配体结合部位的残基,与金属离子结合的残基,形成特定结构motif的残基等等。但并不是所有保守的残基都一定是结构功能重要的,可能它们只是由于历史的原因被保留下来,而不是由于进化压力而保留下来。因此,如果两个序列有显著的保守性,要确定二者具有共同的进化历史,进而认为二者有近似的结构和功能还需要更多实验和信息的支持。通过大量实验和序列比对的分析,一般认为蛋白质的结构和功能比序列具有更大的保守性,因此粗略的说,如果序列之间的相似性超过30%,它们就很可能是同源的。 早期的序列比对是全局的序列比较,但由于蛋白质具有的模块性质,可能由于外显子的交换而产生新蛋白质,因此局部比对会更加合理。通常用打分矩阵描述序列两两比对,两条序列分别作为矩阵的两维,矩阵点是两维上对应两个残基的相似性分数,分数越高则说明两个残基越相似。因此,序列比对问题变成在矩阵里寻找最佳比对路径,目前最有效的方法是Needleman-Wunsch动态规划算法,在此基础上又改良产生了 Smith-Waterman算法和SIM算法。在 FASTA程序包中可以找到用动态规划算法进行序列比对的工具LALIGN,它能给出多个不相互交叉的最佳比对结果。
生物信息学作业 1. Align the leghemoglobin protein from soy bean and myoglobin from human with global and local alignment software (ex. needle and water) respectively and interpret the results. ANSWER: (1)Use Needle to Align the two sequence: Aligned_sequences: 2 # 1: CAA38024.1 # 2: NP_001157488.1 # Matrix: EBLOSUM62 # Gap_penalty: 10.0 # Extend_penalty: 0.5 # Length: 203 # Identity: 43/203 (21.2%) # Similarity: 58/203 (28.6%) # Gaps: 90/203 (44.3%) # Score: 30.0 (2)Use Water to Align the two sequence: Aligned_sequences: 2 # 1: CAA38024.1 # 2: NP_001157488.1 # Matrix: EBLOSUM62 # Gap_penalty: 14 # Extend_penalty: 4 # Length: 32 # Identity: 11/32 (34.4%) # Similarity: 15/32 (46.9%) # Gaps: 0/32 ( 0.0%) # Score: 35 两种软件虽然使用同一罚分标准但得分不同。因为Needle程序实现标准pairwise全局比对,而Water则是局部比对。全局比对因为是比对全长序列,所以空位罚分多,得分较局部比对低。
——古A.名词解释 1. 生物信息学:广义是指从事对基因组研究相关的生物信息的获取,加工,储存,分配,分析和解释。狭义是指综合应用信息科学,数学理论,方法和技术,管理、分析和利用生物分子数据的科学。 2. 基因芯片:将大量已知或未知序列的DNA片段点在固相载体上,通过物理吸附达到固定化(cDNA芯片),也可以在固相表面直接化学合成,得到寡聚核苷酸芯片。再将待研究的样品与芯片杂交,经过计算机扫描和数据处理,进行定性定量的分析。可以反映大量基因在不同组织或同一组织不同发育时期或不同生理条件下的表达调控情况。 3. NCBI:National Center for Biotechnology Information.是隶属于美国国立医学图书馆(NLM)的综合性数据库,提供生物信息学方面的研究和服务。 4. EMBL:European Molecular Biology Laboratory.EBI为其一部分,是综合性数据库,提供生物信息学方面的研究和服务。 5. 简并引物:PCR引物的某一碱基位置有多种可能的多种引物的混合体。 6. 序列比对:为确定两个或多个序列之间的相似性以至于同源性,而将它们按照一定的规律排列。
7. BLAST:Basic Local Alignment Search Tool.是通过比对(alignment)在数据库中寻找和查询序列(query)相似度很高的序列的工具。 8. ORF:Open Reading Frame.由起始密码子开始,到终止密码子结束可以翻译成蛋白质的核酸序列,一个未知的基因,理论上具有6个ORF。 9. 启动子:是RNA聚合酶识别、结合并开始转录所必须的一段DNA序列。原核生物启动子由上游调控元件和核心启动子组成,核心启动子包括-35区(Sextama box)TTGACA,-10区(Pribnow Box)TATAAT,以及+1区。真核生物启动子包括远上游序列和启动子基本元件构成,启动子基本元件包括启动子上游元件(GC岛,CAAT盒),核心启动子(TATA Box,+1区帽子位点)组成。 10. motif:模体,基序,是序列中局部的保守区域,或者是一组序列中共有的一小段序列模式。 11. 分子进化树:通过比较生物大分子序列的差异的数值重建的进化树。 12. 相似性:序列比对过程中用来描述检测序列和目标序列之间相似DNA碱基或氨基酸残基序列所占的比例。 13. 同源性:两个基因或蛋白质序列具有共同祖先的结论。
《生物信息学》上机作业 题目:对人血红蛋白(HBA1)编码基因序列的生物信息分析
目录 引言 .............................................................................................................................................. - 1 -1 正文......................................................................................................................................... - 2 - 1.1 NCBI上对相关核苷酸序列的查找............................................................................ - 2 - 1.2 BLAST运行及其结果.................................................................................................. - 2 - 1.3 BLASTX运行及其结果................................................................................................ - 6 - 2 其他软件的运行及其结果..................................................................................................... - 8 - 2.1 Clustal W运行及其结果 ............................................................................................. - 9 - 2.2 MEGA4.0运行及其结果............................................................................................. - 10 -结论 ............................................................................................................................................ - 10 -
生物信息学分析FAQ CHAPTER ONE ABOUT GENE ONTOLOGY ANNOTATION (3) 什么是GO? (3) GO和KEGG注释之前,为什么要先进行序列比对(BLAST)? (3) GO注释的意义? (3) GO和GOslim的区别 (4) 为什么有些蛋白没有GO注释信息? (4) 为什么GO Level 2的统计饼图里蛋白数目和差异蛋白总数不一致? (4) 什么是差异蛋白的功能富集分析&WHY? (4) GO注释结果文件解析 (5) Sheet TopBlastHits (5) Sheet protein2GO/protein2GOslim (5) Sheet BP/MF/CC (6) Sheet Level2_BP/Level2_MF/Level2_CC (6) CHAPTER TWO ABOUT KEGG PATHWAY ANNOTATION (7) WHY KEGG pathway annotation? (7) KEGG通路注释的方法&流程? (7) KEGG通路注释的意义? (7) 为什么有些蛋白没有KEGG通路注释信息? (8) 什么是差异蛋白的通路富集分析&WHY? (8) KEGG注释结果文件解析 (8) Sheet query2map (8) Sheet map2query (9) Sheet TopMapStat (9) CHAPTER THREE ABOUT FEATURE SELECTION & CLUSTERING (10) WHY Feature Selection? (10)
聚类分析(Clustering) (10) 聚类结果文件解析 (10) CHAPTER FOUR ABOUT PROTEIN-PROTEIN INTERACTION NETWORK (12) 蛋白质相互作用网络分析的意义 (12) 蛋白质相互作用 VS生物学通路? (12) 蛋白质相互作用网络分析结果文件解析 (12)
生物信息学复习题 一、名词解释 生物信息学, 二级数据库, FASTA序列格式, genbank序列格式, Entrez,BLAST,查询序列(query),打分矩阵(scoring matrix),空位(gap),空位罚分,E 值, 低复杂度区域,点矩阵(dot matrix),多序列比对,分子钟,系统发育(phylogeny),进化树的二歧分叉结构,直系同源,旁系同源,外类群,有根树,除权配对算法(UPGMA),邻接法构树,最大简约法构树,最大似然法构树,一致 树(consensus tree),bootstrap,开放阅读框(ORF),密码子偏性(codon bias),基因预测的从头分析法,结构域(domain),超家族,模体(motif),序列表谱(profile),PAM矩阵,BLOSUM,PSI-BLAST,RefSeq,PDB数据库,GenPept, 折叠子,TrEMBL,MMDB,SCOP,PROSITE,Gene Ontology Consortium,表谱(profile)。 二、问答题 1)生物信息学与计算生物学有什么区别与联系? 2)试述生物信息学研究的基本方法。 3)试述生物学与生物信息学的相互关系。 4)美国国家生物技术信息中心(NCBI)的主要工作是什么?请列举3个以上NCBI 维护的数据库。 5)序列的相似性与同源性有什么区别与联系? 6)BLAST套件的blastn、blastp、blastx、tblastn和tblastx子工具的用途 什么? 7)简述BLAST搜索的算法。 8)什么是物种的标记序列? 9)什么是多序列比对过程的三个步骤? 10)简述构建进化树的步骤。 11)简述除权配对法(UPGMA)的算法思想。 12)简述邻接法(NJ)的算法思想。 13)简述最大简约法(MP)的算法思想。 14)简述最大似然法(ML)的算法思想。 15)UPGMA构树法不精确的原因是什么? 16)在MEGA2软件中,提供了多种碱基替换距离模型,试列举其中2种,解释其 含义。 17)试述DNA序列分析的流程及代表性分析工具。 18)如何用BLAST发现新基因? 19)试述SCOP蛋白质分类方案。 20)试述SWISS-PROT中的数据来源。 21)TrEMBL哪两个部分? 22)试述PSI-BLAST 搜索的5个步骤。 三、操作与计算题 1)如何获取访问号为U49845的genbank文件?解释如下genbank文件的LOCUS行提供的信息: LOCUS SCU49845 5028 bp DNA linear PLN 21-JUN-1999 2)利用Entrez检索系统,对核酸数据搜索,输入如下信息,将获得什
CDK2基因和蛋白质序列的生物信息学分析 姓名: 学号: 专业: 1前言 细胞周期蛋白依赖激酶2(cyclin-dependent kinase 2,CDK2),又名细胞分裂激酶2(cell division kinase 2)或p33蛋白激酶(p33 protein kinase),其基因定位于人类基因组的12号染色体上的q13染色带上。CDK2基因全长6013bp,这部分中有7个外显子和6个内含子,7个外显子的长度依次为353bp、78bp、121bp、171bp、102bp、204bp、1264bp(可依次记为外显子1-7)。在翻译过程中,该基因转录成的mRNA的外显子1的前137bp和外显子7的后1159bp不进行翻译,属于调控序列。mRNA上只有中间的部分编码蛋白质。 CDK2基因可以转录为两种mRNA。其中,变体1长度为2325bp,编码298个氨基酸;变体2长度为2223bp,编码264个氨基酸。这两种蛋白质为CDK2的同型蛋白,功能相同,具有调控细胞分裂的功能,主要在G1期到S期和S期到G2期这两个阶段起作用。CDK2广泛分布在生物体的各种细胞的胞质溶胶和细胞核质中,但只在进行分裂的细胞中行使功能,这是因为CDK2只有与不同的细胞周期蛋白(cyclin)结合后才具有活性。CDK2可以与细胞周期蛋白A、B1、B3、E等结合后,参与细胞周期调控。由于CDK2在细胞内的数量变化有可能导致细胞周期异常而产生癌症,故CDK2基因可以被看作癌基因,其活性和表达量可以作为衡量癌症的指标。CDK2与周期蛋白E的复合体不仅能直接参与中心体复制的起始调控,还能与类Rb蛋白p107或转录因子E2F结合,促进细胞从G1期向S期转化或调控DNA复制有关的基因转录。而CDK2与周期蛋白A的复合体可以增强DNA复制因子RF-A的活性。 在CDK2分子中,被称为T环的氨基酸环阻断了活性部位,妨碍激酶履行它的酶功能,而且活性部位的氨基酸形成一种难于为蛋白质结合的形状。CDK2与周期蛋白结合时,周期蛋白将T环转出2nm以上,又将CDK2中的PSTAIRE螺旋部分转了, 并把活性部位氨基酸变成能与底物蛋白结合的正确构象。CDK2的活性不仅与周期蛋白有关,还与其上的Thr-15、Tyr-15、Thr-160三个位点是否磷酸化有关。一般情况下,与周期蛋白结合的CDK2的上述三个位点被Wee/Mik1和CAK激酶磷酸化,但此时复合体还没有活性,只有当Cdc25c将Thr-15、Tyr-15两个位点去磷酸化后,复合体才有活性。细胞中存在多种因子对CDK2进行修饰调节,此外还存在对其活性起负性调控的蛋白质,即CDK激酶抑制物,例如p21CIP/WAF1、p27KIP2等。 前面提到,CDK2基因转录的产物有两种。这两种mRNA的不同之处在于变体1由全部7个外显子组成,而变体2缺失外显子5,由剩余的6个外显子组成。这样翻译成的两种同型蛋白的长度就相差34个氨基酸。 2 材料和方法: 2.1序列数据来源 采用蛋白质名称对NCBI非冗余蛋白质数据库进行检索,CDK2蛋白的记录有1013个。而采用基因名称对NCBI非冗余核酸数据库进行检索,CDK2蛋白的记录有680个。 采用人(Homo sapiens)的CDK2蛋白序列进行BLAST搜索。 2.2序列分析方法
生物信息学分析 生物信息学难吗? 经常有人向我问这个问题,这有什么疑问吗?如果不难学,根本就不用问我这个问题。也无需投入那么多时间精力就能掌握,更无需花费三四千元参加线下的培训班,也不会月薪过万。所以,答案很肯定,道理很简单:生物信息比较难学。 为什么难学? 我总结里几点原因。首先,这是一个交叉学科,要求你既要有生物学的基础,又要有很强的计算机操作技能。这个就有点困难了。因为只是一个生物学就包括多个门类,有很多东西需要去学习,还需要学习计算机知识。很多人一门内容还没学明白,现在还得在加一门,这就属于祸不单行,雪上加霜,屋漏偏逢连夜雨。因此,这种既懂生物学,又懂计算机的复合型人才就比较短缺。而且,生物信息本质上属于数据挖掘,除了生物,计算机,到后面还需要极强的统计学知识才能做好数据分析,所以,还得加上统计学,也就是生物信息学=生物学+计算机科学+统计学三门学科的知识,这也就是为什么生物信息学比较难学。 第二个原因,生物信息本身就包括很多内容,比如DNA的分析,RNA的分析,甲基化的分析,蛋白质的分析等方面,每一
门类又完全不同,从物种方面来分,动物,植物,微生物,医学等有差别很大,很难有一劳永逸,放之四海而皆准的分析方法。 第三个原因就是生物信息是一门快速发展的学习,会出现很多新的测序方法,比如sanger测序,illumina,BGIseq,PacBio,IonTorrent,Nanopore等,每一个平台技术原理完全不同,因此数据特点也完全不同,这就需要针对每一个平台的数据做专门的学习,而且每个平台又在不断的推陈出现,可能今天你刚开发好的方法,产品升级了,都得推倒重来。还有很多新的技术,例如现在比较火的单细胞测序,Hi-C测序,Bionano测序等等内容,以后还出现更多新技术新方法,足够让你活到老,学到老。当然,你先要能活到老,吾生也有涯,而知也无涯。以有涯随无涯,殆已! 高风险才有高收益 当然啦,虽然你已经看到学习生物信息肯定是不容易了,门槛很高,但是呢,门槛高也有很多好处,就是挡住了一部分人,当你学会了,迈过门槛,你的身价就提高了。如果人人都很容易掌握了,那么也就不值钱了。所以,生物信息,前途是光明的,道路是曲折的。
生物信息学中的序列比对算法 张永1,王瑞2 (1.南昌航空大学计算机学院,江西南昌330063;2.江西大宇职业技术学院,江西南昌330038) 摘要:生物信息学是以计算机为工具对生物信息进行储存、检索和分析的科学。序列比对是生物信息学中的一个基本问题,设计快速而有效的序列比对算法是生物信息学研究的一个重要内容,通过序列比较可以发现生物序列中的功能、结构和进化的信息,序列比较的基本操作是比对。本文介绍了序列比对算法的发展现状,描述了常用的各类序列比对算法,并分析了它们的优劣。 关键词:生物信息学;双序列比对;多序列比对 中图分类号:TP301文献标识码:A文章编号:1009-3044(2008)03-10181-04 SequenceAlignmentAlgorithmsinBioinformatics ZHANGYong1,WANGRui2 (1.SchoolofComputing,NanchangHangkongUniversity,Nanchang330063,China;2.JiangxiDayuVocationalInstitute,Nanchang330038,China) Abstract:Bioinformaticsisthesubjectofusingcomputertostore,retrieveandanalyzebiologicalinformation.Sequencealignmentisaba-sicprobleminBioinformatics,anditsmainresearchworkistodeveloprapidandeffectivesequencealignmentalgorithms.Wemaydiscov-erfunctional,structuralandevolutionaryinformationinbiologicalsequencesbysequencecomparing.Thispaperintroducesthedevelop-mentactualityofsequencealignmentalgorithms,describesvarietyofsequencealignmentalgorithmandanalysestheadvantagesanddisad-vantagesofthem. Keywords:Bioinformatics;PairwiseSequenceAlignment;MultipleSequenceAlignment 1引言 生物信息学是80年代末随着人类基因组计划的启动而兴起的一门新的交叉学科,最初常被称为基因组信息学。生物信息学是在生命科学的研究中,以计算机为工具对生物信息进行储存、检索和分析的科学。它是当今生命科学和自然科学的重大前沿领域之一,同时也将是21世纪自然科学的核心领域之一。其研究重点主要体现在基因组学和蛋白组学两方面,具体说,是从核酸和蛋白质序列出发,分析序列中表达结构与功能的生物信息。 生物信息学的研究重点主要体现在基因组学和蛋白质学两方面,具体地说就是从核酸和蛋白质序列出发,分析序列中表达结构和功能的生物信息。生物信息学的基本任务是对各种生物分析序列进行分析,也就是研究新的计算机方法,从大量的序列信息中获取基因结构、功能和进化等知识。在从事分子生物学研究的几乎所有实验室中,对所获得的生物序列进行生物信息学分析已经成为下一步实验之前的一个标准操作。而在序列分析中,将未知序列同已知序列进行相似性比较是一种强有力的研究手段,从序列的片段测定,拼接,基因的表达分析,到RNA和蛋白质的结构功能预测,物种亲缘树的构建都需要进行生物分子序列的相似性比较。例如,有关病毒癌基因与细胞癌基因关系的研究,免疫分子相互识别与作用机制的研究,就大量采用了这类比较分析方法。这种相似性比较分析方法就称为系列比对(SequenceAlignment)。目前,国际互联网上提供了众多的序列比对分析软件。然而,不同的分析软件会得到不同的结果,同时所使用的参数在很大程度上影响到分析的结果。有时常常会由于采用了不合适的参数而丢失了弱的但却具有统计学显著性意义的主要信息,导致随后的实验研究走弯路。因此,生物信息学中的序列比对算法的研究具有非常重要的理论与实践意义。 序列比对问题根据同时进行比对的序列数目分为双序列比对和多序列比对。双序列比对有比较成熟的动态规划算法,而多序列比对目前还没有快速而又十分有效的方法。一般来说,评价生物序列比对算法的标准有两个:一为算法的运算速度,二为获得最佳比对结果的敏感性或准确性。人们虽已提出众多的多序列比对算法,但由于问题自身的计算复杂性,它还尚未得到彻底解决,是 收稿日期:2007-11-25 基金资助:南昌航空大学校自选(EC200706086) 作者简介:张永(1977-),男,硕士,辽宁铁岭人,南昌航空大学计算机学院讲师,研究方向:生物信息学、信息处理;王瑞(1977-),男,江西大宇职业技术学院外语系助教。