
细胞凋亡和细胞增殖都是生命的基本现象,是维持体内细胞数量动态平衡的基本措施。在胚胎发育阶段通过细胞凋亡清除多余的和已完成使命的细胞,保证了胚胎的正常发育;在成年阶段通过细胞凋亡清除衰老和病变的细胞,保证了机体的健康。和细胞增殖一样细胞凋亡也是受基因调控的精确过程,在这一节我们就细胞凋亡的分子机理作简要的介绍。
细胞凋亡的途径主要有两条,一条是通过胞外信号激活细胞内的凋亡酶caspase、一条是通过线粒体释放凋亡酶激活因子激活caspase。这些活化的可将细胞内的重要蛋白降解,引起细胞凋亡。
一、凋亡相关的基因和蛋白
细胞凋亡的调控涉及许多基因,包括一些与细胞增殖有关的原癌基因和抑癌基因。其中研究较多的有ICE、Apaf-1、Bcl-2、Fas/APO-1、c-myc、p53、ATM等。
1.Caspase家族
Caspase属于半胱氨酸蛋白酶,相当于线虫中的ced-3,这些蛋白酶是引起细胞凋亡的关键酶,一旦被信号途径激活,能将细胞内的蛋白质降解,使细胞不可逆的走向死亡。它们均有以下特点:①酶活性依赖于半胱氨酸残基的亲核性;②总是在天冬氨酸之后切断底物,所以命名为caspase(cysteine aspartate-specific protease),方便起见本文称之为凋亡酶;
③都是由两大、两小亚基组成的异四聚体,大、小亚基由同一基因编码,前体被切割后产生两个活性亚基。
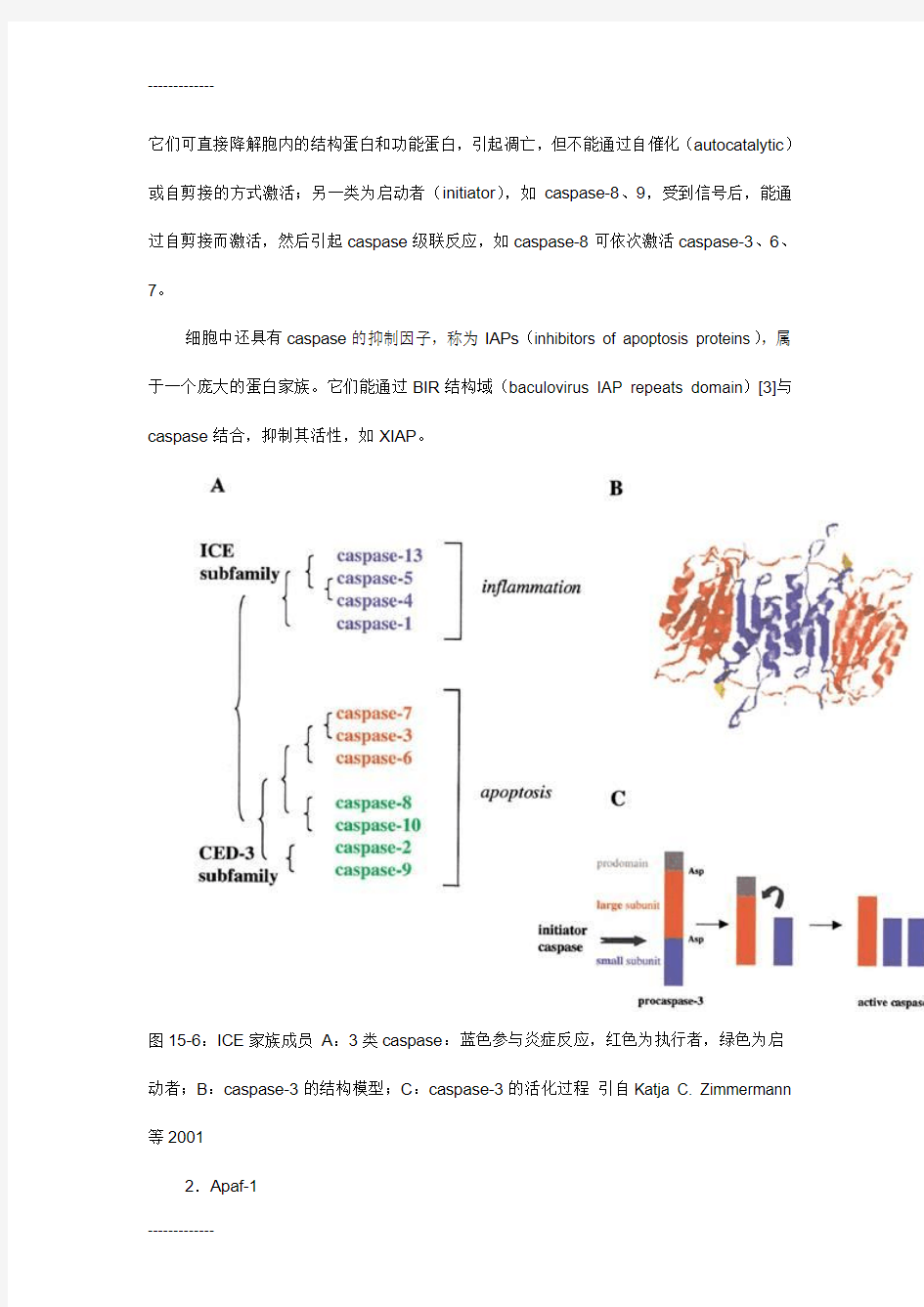
最早发现人类中与线虫ced-3同源的基因[1]是ICE,即:白介素-1 β转换酶(Interleukin-1 β-converting enzyme)基因,因该酶能将白介素前体切割为活性分子,故名。通过cDNA杂交和查找基因组数据库,在人类细胞中已发现11个ICE同源物[2],分为2个亚族(subgroup):ICE亚族和CED-3家族(图15-6),前者参与炎症反应,后者参与细胞凋亡,又分为两类:一类为执行者(executioner或effector),如caspase-3、6、7,
它们可直接降解胞内的结构蛋白和功能蛋白,引起凋亡,但不能通过自催化(autocatalytic)或自剪接的方式激活;另一类为启动者(initiator),如caspase-8、9,受到信号后,能通过自剪接而激活,然后引起caspase级联反应,如caspase-8可依次激活caspase-3、6、7。
细胞中还具有caspase的抑制因子,称为IAPs(inhibitors of apoptosis proteins),属于一个庞大的蛋白家族。它们能通过BIR结构域(baculovirus IAP repeats domain)[3]与caspase结合,抑制其活性,如XIAP。
图15-6:ICE家族成员A:3类caspase:蓝色参与炎症反应,红色为执行者,绿色为启动者;B:caspase-3的结构模型;C:caspase-3的活化过程引自Katja C. Zimmermann 等2001
2.Apaf-1
Apaf-1被称为凋亡酶激活因子-1(apoptotic protease activating factor-1),在线虫中的同源物为ced-4,在线粒体参与的凋亡途径中具有重要作用,该基因敲除后,小鼠神经细胞过多,脑畸形发育。Apaf-1含有3个不同的结构域:①CARD(caspase recruitment domain)结构域,能召集caspase-9;②ced-4 同源结构域,能结合ATP/dATP;③C端结构域,含有色氨酸/天冬氨酸重复序列,当细胞色素c[4]的结合到这一区域后,能引起Apaf-1多聚化而激活。Apaf-1具有激活Caspase-3的作用,而这一过程又需要细胞色素c(Apaf-2)和caspase-9(Apaf-3)参与。Apaf-1/细胞色素c复合体与ATP/dATP结合后,Apaf-1就可以通过其CARD结构域召集caspase-9,形成凋亡体(apoptosome),激活caspase-3,启动caspase级联反应。
3.Bcl-2家族
Bcl-2[5]为凋亡抑制基因,是膜的整合蛋白,其功能相当于线虫中的ced-9。现已发现至少19个同源物,它们在线粒体参与的凋亡途径中起调控作用,能控制线粒体中细胞色素c等凋亡因子的释放。
Bcl-2家族成员都含有1-4个Bcl-2同源结构域(BH1-4),并且通常有一个羧端跨膜结构域(transmembrane region ,TM)。其中BH4是抗凋亡蛋白所特有的结构域,BH3是与促进凋亡有关的结构域。根据功能和结构可将Bcl-2基因家族分为两类(图15-7),一类是抗凋亡的(anti-apoptotic),如:Bcl-2、Bcl-xl、Bcl-w、Mcl-1;一类是促进凋亡的(pro-apoptotic),如:Bax、Bak、Bad、Bid、Bim,在促凋亡蛋白中还有一类仅含BH3结构,如Bid、Bad。
虽然Bcl-2蛋白存在于线粒体膜、内质网膜以及外核膜上,但主要定位于线粒体外膜,它拮抗促凋亡蛋白的功能。而大多数促凋亡蛋白则主要定位于细胞质,一旦细胞受到凋亡因子的诱导,它们可以向线粒体转位,通过寡聚化在线粒体外膜形成跨膜通道,或者开启线粒体的PT孔,从而导致线粒体中的凋亡因子释放,激活caspase,导致细胞凋亡。
胞质中的促凋亡蛋白可通过不同的方式被激活,包括去磷酸化,如Bad;被caspase 加工为活性分子,如Bid;从结合蛋白上释放出来,如Bim是与微管蛋白结合在一起的。
图15-7 Bcl-2家族引自Katja C. Zimmermann等2001 4.Fas
Fas又称作APO-1/CD95,属TNF受体家族。Fas基因编码产物为分子量45KD的跨膜蛋白,分布于胸腺细胞,激活的T和B淋巴细胞,巨噬细胞,肝、脾、肺、心、脑、肠、睾丸和卵巢细胞等。Fas蛋白与Fas配体结合后,会激活caspase,导致靶细胞走向凋亡。
5.p53
是一种抑癌基因,其生物学功能是在G期监视DNA的完整性。如有损伤,则抑制细胞增殖,直到DNA修复完成。如果DNA不能被修复,则诱导其调亡,研究发现丧失p53功能的小鼠胸腺细胞对糖皮质激素诱导的调亡反应和正常细胞相同,而对辐射诱导的调亡不
敏感。
6.myc
在许多人类恶性肿瘤细胞中都发现有c-myc的过度表达,它能促进细胞增殖、抑制分化。在凋亡细胞中c-myc也是高表达,作为转录调控因子,一方面它能激活那些控制细胞增殖的基因,另一方面也激活促进细胞凋亡的基因,给细胞两种选择:增殖或凋亡。当生长因子存在,Bcl-2基因表达时,促进细胞增殖,反之细胞凋亡。
7.ATM
ATM(ataxia telangiectasia-mutated gene)是与DNA损伤检验有关的一个重要基因。最早发现于毛细血管扩张性共济失调症患者,人类中大约有1%的人是ATM缺失的杂合子,表现出对电离辐射敏感和易患癌症。正常细胞经放射处理后,DNA损伤会激活修复机制,如DNA不能修复则诱导细胞凋亡。ATM是DNA损伤检验点的一个重要的蛋白激酶(参见第十三章第四节)
二、Fas介导的细胞凋亡
细胞表面的凋亡受体是属于肿瘤坏死因子受体(TNFR)家族的跨膜蛋白,它们包括Fas(Apo-1/CD95)、TNFR1、DR3/WSL、DR4/TRAIL-R1和DR5/TRAIL-R2。其配体属于TNF家族,目前已比较清楚的是Fas介导的细胞凋亡途径。
Fas具有三个富含半胱氨酸的胞外区和一个称为死亡结构域(Death domain,DD,图15-8)的胞内区。Fas的配体FasL(Fas ligand)与Fas结合后,Fas三聚化使胞内的DD 区构象改变,然后与接头蛋白FADD(Fasassociated death domain)的DD区结合,而后
FADD的N端DED区(death effector domain)就能与Caspase-8(或-10)前体蛋白结合,形成DISC (death-inducing signaling complex )[6] ,引起caspase-8、10通过自身剪激活,它们启动caspase的级联反应,使caspase-3、-6、-7激活,这几种Caspase可降解胞内结构蛋白和功能蛋白,最终导致细胞凋亡。
图15-8 FAS介导的细胞凋亡引自Avi Ashkenazi and Vishva M. Dixit 1998 Caspase 可激活名叫CAD(caspase-activated Dnase)的核酸酶,CAD能在核小体的连接区将其切断,形成约为200bp整数倍的核酸片段。正常情况下CAD存在于胞质中,
并且与抑制因子ICAD/DFF-45蛋白结合,不能进入细胞核。Caspase活化后可以降解ICAD/DFF-45,释放出CAD,使它进入细胞核降解DNA。
Fas/FasL系统在免疫系统中具有重要的作用,其一是参与免疫调节,活化成熟的外周T细胞主要通过Fas/FasL系统介导的细胞凋亡清除与自身抗原有交叉反应的克隆和由自身抗原激活的细胞克隆,以限制T细胞克隆的无限增殖,防止对自身组织的损伤,即产生外周免疫耐受。淋巴细胞凋亡异常导致的免疫耐受失控,是自身免疫性疾病的主要病因。其二是细胞毒T细胞(CTL)可以通过FasL诱导靶细胞凋亡,但遗憾的是,某些肿瘤细胞也可以通过这一途径诱导淋巴细胞凋亡,从而逃脱免疫监控。
三、线粒体与细胞凋亡
细胞应激反应或凋亡信号能引起线粒体细胞色素c释放,作为凋亡诱导因子,细胞色素c能与Apaf-1、caspase-9前体、ATP/dATP形成凋亡体(apoptosome,图15-9),然后召集并激活caspase-3,进而引发caspases级联反应,导致细胞凋亡。
在这里,一个核心的问题是细胞色素c究竟通过哪一种途径释放到细胞质中,由于大部分凋亡细胞中很少发生线粒体肿胀和线粒体外膜破裂的现象,所以目前普遍认为细胞色素是通过线粒体PT孔或Bcl-2家族成员形成的线粒体跨膜通道释放到细胞质中的。
线粒体PT孔(permeability transition pore)主要由位于内膜的腺苷转位因子(Adenine nucleotide translocator,ANT)和位于外膜的电压依赖性阴离子通道(Voltage dependent anion channel,VDAC)等蛋白所组成,PT孔开放会引起线粒体跨膜电位下降和细胞色素c释放。Bcl-2家族蛋白对于PT孔的开放和关闭起关键的调节作用,促凋亡蛋白Bax等可以通过与ANT或VDAC的结合介导PT孔的开放,而抗凋亡类蛋白如Bcl-2、Bcl-xL等则
可通过与Bax竞争性地与ANT结合,或者直接阻止Bax与ANT、VDAC的结合来发挥其抗凋亡效应。
Bcl-2家族的结构和能形成离子通道的一些毒素(如大肠杆菌毒素)非常相似。插入膜结构中形成较大的通道,允许细胞色素c等蛋白质通过,这可能是细胞色素c释放的另一个途径。
在线虫中ced-3和ced-4的缺失突变抑制所有发育阶段的细胞死亡。在哺乳动物中,尽管Apaf-1基因缺失的小鼠没有caspase活化,但除了神经细胞过多外,大多数器官发育是正常的。近年来的研究发现随细胞色素c释放的蛋白还有Smac(second mitochondria-derived activator of caspase)、凋亡诱导因子(apoptosis inducing factor,AIF)和核酸内切酶G( Endo G)。Smac能通过N端的几个氨基酸与IAPs(凋亡抑制蛋白)的BIR结构域结合,从而解除IAP对caspase的抑制;AIF[7]则引起核固缩和染色质断裂;Endo G可以使DNA片段化。可见即使在caspase不参与的情况下,由线粒体途径仍可引起细胞凋亡。
在对Fas应答的细胞中,一型细胞(type I),如胸腺细胞,其caspase-8有足够的活性,被Fas活化后导致细胞凋亡,在这类细胞中高表达Bcl-2不能抑制Fas诱导的细胞凋亡。在二型细胞(type II),如肝细胞中,Fas介导的caspase-8活化不能达到足够的水平,因此这类细胞中的凋亡信号需要借助凋亡的线粒体途径来放大。活化的caspase-8将胞质中的Bid剪切,形成活性分子tBid(truncated Bid),tBid进入线粒体,导致细胞色素c释放,使凋亡信号放大。
图15-9 细胞色素释放引起的凋亡引自R. Chris Bleackley and Jeffrey A. Heibein 2001
我们不看出线粒体既是细胞的能量工厂,也是细胞的凋亡控制中心,可是为什么线粒体会担负起如此重要的双重功能呢?一个主要的原因是各类生长因子都可以促进葡萄糖转运和己糖激酶等向线粒体转运、加速能量生产,相反地剥夺生长因子后,细胞氧消耗降低、ATP合成不足、蛋白质合成受阻,最后细胞走向死亡。由于这一方面的资料较少,目前还很难作出一个较好的解释,只能留在以后再完善。
[1]Horvitz实验室的袁均英1993年发现哺乳动物ced-3的同源物为白介素-1-β转换酶(ICE)。
[2]哺乳动物中已发现14个。
[3]最早在细菌和病毒中发现。
[4]是线粒体内膜的外周蛋白,呼吸链中的两个可移动组分之一,位于膜间隙,释放到细胞质中会引起细胞凋亡。
[5] 是一种原癌基因,名称来源于B细胞淋巴瘤/白血病-2(B-cell lymphoma/Leukemia-2,bcl-2),最早由Tsujimoto(1985)从伴有14、18染色体易位的淋巴瘤细胞中发现,在正常人体内位于18号染色体,在患者易位于14号染色体。
[6] Kischkel等1995发现Fas活化时可以与至少4种蛋白相连,分别称为CAP1(Cytotoxicity-dependent APO-1 (Fas/CD95)-associated proteins 1)、CAP2、CAP3和CAP4,这4种蛋白与活化的Fas受体一起被称为死亡诱导信号复合物(death-inducing signaling complex, DISC)。随后的研究证实CAP1和CAP2是不同形式丝氨酸磷酸化的FADD蛋白,CAP3和CAP4实际上就是活化的caspase-8。
[7]是一种依赖于黄素的一种氧化还原酶,目前还不清楚其作用机制。
细胞凋亡和细胞增殖都是生命的基本现象,是维持体内细胞数量动态平衡的基本措施。在胚胎发育阶段通过细胞凋亡清除多余的和已完成使命的细胞,保证了胚胎的正常发育;在成年阶段通过细胞凋亡清除衰老和病变的细胞,保证了机体的健康。和细胞增殖一样细胞凋亡也是受基因调控的精确过程,在这一节我们就细胞凋亡的分子机理作简要的介绍。 细胞凋亡的途径主要有两条,一条是通过胞外信号激活细胞内的凋亡酶caspase、一条是通过线粒体释放凋亡酶激活因子激活caspase。这些活化的可将细胞内的重要蛋白降解,引起细胞凋亡。 一、凋亡相关的基因和蛋白 细胞凋亡的调控涉及许多基因,包括一些与细胞增殖有关的原癌基因和抑癌基因。其中研究较多的有ICE、Apaf-1、Bcl-2、Fas/APO-1、c-myc、p53、ATM等。 1.Caspase家族 Caspase属于半胱氨酸蛋白酶,相当于线虫中的ced-3,这些蛋白酶是引起细胞凋亡的关键酶,一旦被信号途径激活,能将细胞内的蛋白质降解,使细胞不可逆的走向死亡。它们均有以下特点:①酶活性依赖于半胱氨酸残基的亲核性;②总是在天冬氨酸之后切断底物,所以命名为caspase(cysteine aspartate-specific protease),方便起见本文称之为凋亡酶; ③都是由两大、两小亚基组成的异四聚体,大、小亚基由同一基因编码,前体被切割后产生两个活性亚基。 最早发现人类中与线虫ced-3同源的基因[1]是ICE,即:白介素-1 β转换酶(Interleukin-1 β-converting enzyme)基因,因该酶能将白介素前体切割为活性分子,故名。通过cDNA杂交和查找基因组数据库,在人类细胞中已发现11个ICE同源物[2],分为2个亚族(subgroup):ICE亚族和CED-3家族(图15-6),前者参与炎症反应,后者参与细胞凋亡,又分为两类:一类为执行者(executioner或effector),如caspase-3、6、7,
凋亡相关基因Bax、Bad和Bcl-2与PTSD的相关性 肖甄男 中国医科大学 93期 【摘要】创伤后应激障碍(PTSD)是指由于异常威胁性或灾难性心理创伤导致延迟出现和长期持续的精神障碍。PTSD患者有强烈的恐惧和惊吓反应,血中糖皮质激素浓度反常低下,下丘脑一垂体一肾上腺轴(HPA)轴调节紊乱,激活糖皮质激素受体进而上调促凋亡基因Bax,Bad引起海马神经元凋亡,海马萎缩,体积缩小。从而影响记忆学习功能的发挥,造成不同的记忆损害甚至导致记忆缺失。杏仁核是大脑中的“恐惧中枢”,杏仁核和海马的功能密切相关,海马的功能受杏仁核的调节而实现。 PTSD导致凋亡相关基因Bax和Bcl一2比值增大,使杏仁核神经元细胞出现凋亡,杏仁核变稀疏,功能减退。Bcl-2相关家族在海马,杏仁核细胞凋亡中起到了关键性的作用 The Relationship of Apoptosis—related Genes Bax,Bad and Bcl-2 with Pathogenesis of Post—traumatic Stress Disorder XIAO Zhen-nan (93k, China Medical University) 【Abstract】 Post-traumatic stress disorder(PTSD)is an anxiety disorder that can develop after exposure to one or more traumatic events threatened or caused grave physical harm. PTSD patients have a strong sense of fear and startle response,abnormal low blood glucocorticoid concentration,disordered HPA axis, activation of glucocorticoid receptor and the expression of apoptosis—related genes Bax,Bad is down-regulated. These changes cause neuronal apoptosis in hippocampus, hippocampus atrophy,volume decreased and affect the exertion of memory and learning function,inducing different memory impairment even memory lost. Amygdala nucleus is the brain’s “fear center”.The function of hippocampus is closely linked with amygdala and realized by the regulation of amygdala. PTSD increases the ratio of Bax/ Bc1—2 and induces neuronal apoptosis in amygdala, causing the amygdala sparse and hypofunction. Bcl-2 related family play an important role in the neuronal apoptosis in hippocampus and amygdala. 创伤后应激障碍(posttraumatic stress disorder,PTSD)是指由于灾害、战争、恐怖事件、交通意外等引起的巨大痛苦或受惊吓、遭受悲剧导致的精神、身心症状的持续状态或焦虑综合征。是应激疾病中最为典型的一种[1],严重影响人们的生存质量。PTSD患者有强烈的恐惧和惊吓反应;其血中糖皮质激素浓度反常低下,HPA 轴调节紊乱。在严重创伤应激急性期,在中枢神经系统(CNS)皮质边缘区,特别是杏仁核、海马和下丘脑等处短期内神经可塑性改变,提示这些脑区可能为CNS应激应答敏感区。研究表明,PTSD大鼠海马萎缩[2],海马神经元发生凋亡[3],杏仁核和海马的功能密切相关,海马的部分功能可由杏仁核的调节而实现[4],杏仁核作为边缘系统中重要的皮质下核团,在PTSD时也发生细胞凋亡[5]。杏仁核与海马神经元细胞的凋亡可能与PTSD的发病密切相关。 1. 细胞凋亡 细胞凋亡是指为维持内环境稳定而由基因控制的,机体正常细胞在受到生理和病理性刺激后出现的一种自发的,在自身基因调控下进行的一种主动死亡,是机体维持自身稳定的一种基本生理机制。在生理条件下细胞凋亡主要是作为机体调节细胞群生长消亡平衡的重要手段,与细胞增殖一起维持机体细胞群的自身稳定。细胞凋亡与坏死有本质的不同,凋亡是一
万方数据
凋亡抑制蛋白家族的研究进展 作者:高艳霞, 刘仲娟 作者单位:南昌大学第一附属医院耳鼻喉科,江西南昌,3300066 刊名: 中外医疗 英文刊名:CHINA FOREIGN MEDICAL TREATMENT 年,卷(期):2010,29(25) 被引用次数:0次 参考文献(4条) 1.Hay BA UnderStanding IAP function and regulation:a view from Drosophila 2000(11) 2.Joazeiro CA.Weissman AM RING finger proteins:mediators of ubiquitin ligase activity 2000(5) 3.Roy N.Deveraux QL.Takashashi R The c-IAP-1 and c-IAP-2 proteins are direct inhibitors of specific caspases 1997(23) 4.Deveraux QL.Roy N.Stennicke HR IAPs block apoptosis events induced by caspase-8 and cytochrome c by direct inhibition of distinct caspases 1998(8) 本文链接:https://www.doczj.com/doc/1512736669.html,/Periodical_hgzy201025152.aspx 授权使用:云南大学(yndx),授权号:ac529863-c7c4-4fda-94bd-9e6901883986 下载时间:2011年1月12日
X连锁凋亡抑制蛋白与肝细胞肝癌的研究进展X连锁凋亡抑制蛋白(X-linked inhibitor of apoptosis protein,XIAP,又称 human IAP-like protein,hILP)是凋亡抑制蛋白家族(inhibitor of apoptosis proteins,IAPs)中的其中一个重要成員。IAPs首先在杆状病毒中发现,能抑制感染病毒 的宿主细胞凋亡,随后逐渐发现在人类及其他物种中广泛存在。XIAP与杆状病毒IAPs有高度同源性,均具有显著的抗凋亡作用。 1 XIAP的结构特点 人类XIAP基因定位于Xq25,首先由Duckett克隆其全长cDNA,含2540bp。XIAP由497个氨基酸组成,分子量为57KD,N端有3个IAP重复序列BIR1、BIR2和BIR3,与抑制细胞凋亡相关,C端有RING锌指样结构域,具有泛素连接酶E3活性,可自身泛素化及泛素化其他底物,从而也调节细胞的凋亡[1]。另外,在XIAP的BIR和RING之间,存在一个泛素结合区域(UBA),与NF-κB 的激活有关[2]。 2 XIAP的抗凋亡作用主要途径 2.1抑制caspase caspase是细胞凋亡的执行者。正常情况下以无活性酶原形成存在,在凋亡信号传递下发生瀑布式激活,最终导致细胞凋亡。Deveraux[3]发现,XIAP可直接抑制caspase3、7的活性,而caspase3位于caspase凋亡通路的下游,为caspase凋亡通路中的效应蛋白,为凋亡的执行者。XIAP通过N端的BIR2与caspase3、7的活性部位结合,阻止caspase3、7与底物的结合和进一步级联催化反应[4]。Shiozaki指出,XIAP的BIR3结构域与caspase9单体结合形成异源二聚体,使caspase9保持单体结构,失去催化活性,不能启动caspase 级联反应。caspase9位于caspase凋亡途径的上游[5]。因而XIAP可同时作用于caspase的起始阶段及效应阶段。另外,XIAP的C端RING环,可以通过泛素化导致caspase的降解,同样达到抑制caspase途径的凋亡。 2.2 JNK1信号通路JNK1是丝裂原活化蛋白激酶MAPK重要通路之一,是XIAP另外一条主要抗凋亡途径。XIAP抗凋亡机理中,JNK1信号通路在不同细胞中的作用尚有争论。在IL-1β转换酶诱发的凋亡过程中,XIAP选择性激活JNK,从而达到抑制凋亡的作用[6]。XIAP的过表达激活JNK1信号通路,也可阻止TNF-α,ICE、FAS-诱导的293、MCF7细胞凋亡[7]。ILPIP(人类ILP交互作用蛋白)可以增强XIAP对JNK1的激活,抑制由TNF-α介导的凋亡[8]。而Kaur 对鼠肝细胞的研究中发现,XIAP促进泛素化降解TAK1(transformin growth facter-β-activated kinase),阻断JNK1信号通路,从而抑制由TGF-β1介导的细胞凋亡[9]。 2.3通过NF-κB途径NF-κB是一种重要转录调节因子。NF-κB信号转导通路与肿瘤的发生、增殖、分化、抗凋亡、侵袭和转移有密切关系。肝细胞癌变过程中NF-κB信号转导途径激活和Bcl-2呈过表达状态与肝癌的发生、发展密切相
细胞凋亡及相关因素的研究进展 论文摘要:细胞凋亡(Apoptosis)是一种生理性死亡Physiogicalcell death,PCD),是细胞对内外信息刺激的 应答反应,[1]它与细胞的生长、分化一样.属于最基本的 细胞学事件或过程.它决定着生物体的基本特征和转归.是胚胎发生和个体发育中清除细胞以维持细胞数目正常的调 节机制。 [2]当组织细胞发生异常调亡时,即可引起疾病的发生。一般来讲.凋亡过多会引起退行性变或早衰,调亡过少.易诱发肿瘤。[3] 因此,细胞凋亡近年来引起生命科学研究领域的广泛关注。本文仅就细胞调亡的概念及相关因素作一简要的概述。 【Summary】 Apoptosis is a kind of Physiogicalcell death and a reaction of cells to around informations stimulate .[1] It is same as cells’ growth and differentiation which belong to the basic cell subject’s incident or process.it decide living things’essential character- Istics ,and used to clear away cells to keep it rgular number’s regulation mechanism in the procees of embryo occur and individual growth.[2]there will couse ill- Ness when the organization cells come into being particularly apoptosis .Generally speaking ,more cell
细胞凋亡,即程序性细胞死亡,在维持多细胞组织生长和保持体内细胞动态平衡方面发挥着重要作用。如果细胞凋亡发生紊乱,平衡将被打乱进而导致各种疾病。细胞凋亡存在两条途径:内源性途径和外源性途径。内源性途径需要线粒体参与细胞凋亡信号的放大来调节细胞凋亡,而外源性途径不需要线粒体的参与,而是直接激活细胞膜上死亡受体诱导细胞凋亡[1]。在肿瘤细胞中,细胞很少或几乎不凋亡,出现此种凋亡异常现象多是由于促凋亡因子的下调和(或)细胞内 [文章编号]1007-7669(2013)04-0257-08 凋亡抑制蛋白及其拮抗剂研发进展 范应仙a,张继虹a,b,张宽仁a (昆明理工大学a.生命科学与技术学院,b.医学院,云南昆明650500) [关键词]细胞凋亡;凋亡抑制蛋白质类;smac蛋白;拟态物;肿瘤治疗方案 [摘要]凋亡抑制蛋白(IAPs)是一类重要的细胞内凋亡调节蛋白,与肿瘤细胞的抗凋亡能力及耐药性密切相关。线粒体释放的蛋白smac是一种抗凋亡拮抗蛋白,通过其N端四肽序列(Ala-Val-Pro-Ile,AVPI)与IAPs的杆状病毒IAP重复序列(BIRs)结合,从而释放半胱氨酸天冬氨酸蛋白酶(caspases),促进细胞的凋亡。近年来以IAPs为靶点,研制有抗癌特异性的小分子IAP拮抗剂,寻找高效低毒的smac 拟态物(smac-mimics)的治疗策略倍受关注。Smac-mimics的研发是近几年研究用于肿瘤治疗的一种新思路。本文主要介绍IAPs作用机制及抗凋亡拮抗剂smac-mimics研发进展。 [中图分类号]R966[文献标志码]A Development of inhibitors of apoptosis proteins and its antagonist FAN Ying-xian a,ZHANG Ji-hong a,b,CHANG Kwen-jen a (a.Faculty of Life Science and Technology,b.Faculty of Medicine,Kunming University Science and Technology,Kunming YUNNAN650500,China) [KEY WORDS]apoptosis;inhibitor of apoptosis proteins;smac protein;mimic;antineoplastic protocols [ABSTRACT]Inhibitors of apoptosis proteins(IAPs)are potent inhibitors of apoptosis by binding caspases and inhibiting their enzymatic activities,which are associated with the tumour anti-apoptosis and resistance.The protein second-mitochondria derived activator of caspases(smac),an antagonist of IAPs,released from mitochondria can also bind to IAPs through its N-terminal AVPI(Ala-Val-Pro-Ile)tetrapeptide and releases active caspases from IAPs,and thus reactivates apoptosis.Several smac mimetic based on AVPI structure were discovered recently as potent antagonists of IAPs,and became a new strategy for cancer therapy.This paper mainly introduced the action mechanism of IAPs and research progress of smac mimetic. [收稿日期]2011-11-23[接受日期]2012-10-30 [基金项目]云南省自然科学基金(KKSA201126061) [作者简介]范应仙,女,硕士研究生,主要从事分子药理学研究,E-mail:19861111fyx@https://www.doczj.com/doc/1512736669.html, [责任作者]张继虹,E-mail:zhjihong2000@https://www.doczj.com/doc/1512736669.html,
生物化学与生物 物理进展 PROGRESS IN BIOCHCMISTRY AND BIOPHYSICS 1999年 第1期 No.1 1999 TF-1细胞凋亡相关基因的研究 刘红涛 王玉刚 张颖妹 宋泉声 敬保迁 袁 勇 马大龙 摘要 利用近年来发展起来的代表差异分析(cDNA representational differences analysis, cDNA-RDA)技术研究了在人红白血病细胞株TF-1细胞撤除细胞因子后进入凋亡时诱导表达的基因.发现了6个新基因片段.其中有三个经与GenBank nr和dbEST查询均没有发现同源性,已经向GenBank进行登记,登记号分别为U83208,U83279, U83397.此外还发现一批已知基因的表达与凋亡相关,其中包括Hou和人硫氧还原蛋白等, 提示它们在凋亡中可能起作用.这项工作为进一步研究凋亡相关基因打下了良好基础.通过RDA的研究结果,有可能发现人白血病细胞凋亡的特异标记蛋白或发挥作用的重要蛋白,以期为白血病治疗提供理论基础. 关键词 TF-1细胞株,代表差异分析,凋亡 学科分类号 R392.1 Studies on the Apoptosis-Related Genes of TF-1 Cell Line by cDNA-RDA Technique. LIU Hong-Tao, WANG Yu-Gang, ZHANG Ying-Mei, SONG Quan-Sheng, JING Bao-Qian, YUAN Yong, MA Da-Long (Department of Immunology, Beijing Medical University, Beijing 100083, China). Abstract The genes effecting in the process of the apoptosis of TF-1 cell line when it is deprived of the cytokine in the culture medium were studied by RDA (representational difference analysis) method. The TF-1 cell depriving of cytokines for 8 hours was selected as the Tester and normal-cultured TF-1 cell as the Driver. Seven gene fragments were found uniquely expressed or highly expressed in the process of apoptosis of TF-1 cell line which include some known genes such as Hou and thioredoxin that formerly suggested to play a role in apoptosis.There are three fragments are complete novel after searching the nr and EST catalogues of GenBank and were banked into GenBank. The accession numbers for them are U83208, U83279, U83397 respectively. From the novel gene fragments, the complete cDNA sequence of them can be fished and the bioactivity and function of them in apoptosis of TF-1 cell and other hematological tumors can be further studied. On the other hand, the function of some known genes which were not suggested formerly in the course of apoptosis can be studied. Key words TF-1 cell line, representational difference analysis(RDA), apoptosis
凋亡相关基因BaxBad和Bcl
————————————————————————————————作者:————————————————————————————————日期:
凋亡相关基因Bax、Bad和Bcl-2与PTSD的相关性 肖甄男 中国医科大学 93期 【摘要】创伤后应激障碍(PTSD)是指由于异常威胁性或灾难性心理创伤导致延迟出现和长期持续的精神障碍。PTSD患者有强烈的恐惧和惊吓反应,血中糖皮质激素浓度反常低下,下丘脑一垂体一肾上腺轴(HPA)轴调节紊乱,激活糖皮质激素受体进而上调促凋亡基因Bax,Bad引起海马神经元凋亡,海马萎缩,体积缩小。从而影响记忆学习功能的发挥,造成不同的记忆损害甚至导致记忆缺失。杏仁核是大脑中的“恐惧中枢”,杏仁核和海马的功能密切相关,海马的功能受杏仁核的调节而实现。 PTSD导致凋亡相关基因Bax和Bcl一2比值增大,使杏仁核神经元细胞出现凋亡,杏仁核变稀疏,功能减退。Bcl-2相关家族在海马,杏仁核细胞凋亡中起到了关键性的作用 The Relationship of Apoptosis—related Genes Bax,Bad and Bcl-2 with Pathogenesis of Post—traumatic Stress Disorder XIAO Zhen-nan (93k, China Medical University) 【Abstract】 Post-traumatic stress disorder(PTSD)is an anxiety disorder that can develop after exposure to one or more traumatic events threatened or caused grave physical harm. PTSD patients have a strong sense of fear and startle response,abnormal low blood glucocorticoid concentration,disordered HPA axis, activation of glucocorticoid receptor and the expression of apoptosis—related genes Bax,Bad is down-regulated. These changes cause neuronal apoptosis in hippocampus, hippocampus atrophy,volume decreased and affect the exertion of memory and learning function,inducing different memory impairment even memory lost. Amygdala nucleus is the brain’s “fear center”.The function of hippocampus is closely linked with amygdala and realized by the regulation of amygdala. PTSD increases the ratio of Bax/ Bc1—2 and induces neuronal apoptosis in amygdala, causing the amygdala sparse and hypofunction. Bcl-2 related family play an important role in the neuronal apoptosis in hippocampus and amygdala. 创伤后应激障碍(posttraumatic stress disorder,PTSD)是指由于灾害、战争、恐怖事件、交通意外等引起的巨大痛苦或受惊吓、遭受悲剧导致的精神、身心症状的持续状态或焦虑综合征。是应激疾病中最为典型的一种[1],严重影响人们的生存质量。PTSD患者有强烈的恐惧和惊吓反应;其血中糖皮质激素浓度反常低下,HPA 轴调节紊乱。在严重创伤应激急性期,在中枢神经系统(CNS)皮质边缘区,特别是杏仁核、海马和下丘脑等处短期内神经可塑性改变,提示这些脑区可能为CNS应激应答敏感区。研究表明,PTSD大鼠海马萎缩[2],海马神经元发生凋亡[3],杏仁核和海马的功能密切相关,海马的部分功能可由杏仁核的调节而实现[4],杏仁核作为边缘系统中重要的皮质下核团,在PTSD时也发生细胞凋亡[5]。杏仁核与海马神经元细胞的凋亡可能与PTSD的发病密切相关。 1. 细胞凋亡 细胞凋亡是指为维持内环境稳定而由基因控制的,机体正常细胞在受到生理和病理性刺激后出现的一种自发的,在自身基因调控下进行的一种主动死亡,是机体维持自身稳定的一种基本生理机制。在生理条件下细胞凋亡主要是作为机体调节细胞群生长消亡平衡的重要手段,与细胞增殖一起维持机体细胞群的自身稳定。细胞凋亡与坏死有本质的不同,凋亡是一
细胞凋亡的基因调控机制近年来,对线虫的研究发现存在十余个与细胞 凋亡有关的基因,与后来在哺乳动物中找到的相关 基因结构和功能相似,是同源基因,说明细胞凋亡 基因在动物进化的过程中高度保守。目前发现三类 与细胞凋亡有关的基因:促细胞凋亡基因、抑制细胞 凋亡基因和在细胞凋亡过程中协助的基因。包括: ①ced基因:线虫的eed.3和ced一4为促凋亡基因, 哺乳类动物和eed.3相似基因为ICE,是一种半胱 氨酸蛋白酶;ted一9是抑制凋亡基因Il9】,人的bcl-2和物促凋亡基因。ICE作用于IL.1p 前体。抑制ICE的 基因有牛痘病毒的Crm A和杆状病毒的P53,对细 胞凋亡都有较强的抑制作用;③hd.2家族:hc1.2家 族有Bcl—x、Bax、Bak和Bad等成员,bc1.2可以通过 CED一4作用调节caspases和线粒体通透性转变,从 而防止凋亡产生,与肿瘤细胞增殖和癌变有关[201,而 Bax有促进凋亡的作用;④c.myc家族:c.myc具有促 进细胞增殖和促使细胞凋亡双向的调节作用,这主 要决定于其上游的信号来源;~TNV受体家族:Fas 是TNF受体家族中研究最详细的成员,Fas为死亡 受体。Fas—L是T淋巴细胞产物,Fas和Fas.L结合诱 导细胞凋亡;~p53基因:野生型诱导细胞凋亡,通 过细胞凋亡抑制肿瘤的生长。它作用在细胞周期 和细胞受损伤时,促使细胞凋亡;⑦Rh和E2F:Rb 有促凋亡作用,E2F包括E2F一1、E2F一2、E2F一3、E2F一4 和E2F一5,共5种紧密相关的转录因子,其中E2F一1、 E2F,2和E2F,3结合Rb蛋白,而E2F一4和E2F-5主 要结合Rb的相关蛋白p107和p130。E2F一1具有双 重作用,当E2F一1与Rh蛋白结合时,细胞周期进程 被抑制。但当Rh蛋白缺失时E2F一1刺激细胞增殖。 可能细胞内E2F的浓度或者其它细胞周期调节因 子存在与否,影响E2F.1决定发挥细胞生长还是细 胞凋亡作用[Z21;⑧ras:ras是癌基因,它的作用是抑制 细胞凋亡。
?综述? PKCδ诱导的细胞凋亡相关蛋白研究进展3 张炜,叶华,叶菜英△,张德昌 【摘要】蛋白激酶C是一个庞大的家族,参与了细胞内多种信息传导通路,具有复杂的生理功能,目前认为其亚型之一PKCδ可诱导细胞的凋亡。该过程信号通路复杂,相关蛋白种类繁多,除常见的caspase家族以外,p53、PLS-3、拓扑异构酶、Mcl-1等蛋白也参与了PKCδ对凋亡的调节过程。 【关键词】PKCδ;凋亡;p53;PLS-3 【中图分类号】R364 【文献标识码】 A 【文章编号】1606-8106(2007)02-0128-03 蛋白激酶C(PKC)是一类由多种同工酶组成的丝/苏氨酸蛋白激酶家族,处于磷脂酰肌醇代谢应答细胞所受到刺激的中心环节,与神经传导、肿瘤生长及细胞的生长、分化和凋亡等生理病理过程有着密切的联系。其中,PKCδ,一种不依赖于Ca2+的nPKC亚型,现认为可促进细胞凋亡过程[1]。在各种刺激因素的作用下,PKCδ可移位至线粒体[2]或细胞膜,促使caspase3活化[3],将其切割并释放出活性片断[4],进而磷酸化多种底物蛋白,使之活化或失活,最终引起细胞凋亡。在这个过程中,有多种细胞因子如细胞色素c,c-Abl, AnnexinV[5]NGF/p75[6]DNA-PK等都参与了细胞凋亡的调节,本文只就其中一部分,p53、PLS-3、拓扑异构酶Ⅱα和Mcl-1在PKCδ依赖的细胞凋亡途径中所起作用做一个简单的阐述。 1DNA结合蛋白p53 p53,是一种DNA结合蛋白,由p53基因编码的细胞核磷蛋白,作为转录因子,可调节多种基因的表达,如p21、Bax、Bcl-2、Gadd等,并可以与多种蛋白如c-Abl等相互作用[7]。其正常功能为调控细胞分化,维持细胞基因的完整性,对DNA损伤的修复以及细胞周期的正常运转。白血病、骨肉瘤、肺癌、直肠结肠癌中,p53蛋白发生突变或缺失。 对于P53在细胞死亡信号过程中的作用,已经进行了很多的研究。活性氧或者活性氮处理细胞,可引起细胞死亡,其中细胞内p53蛋白水平升 3基金项目:“973”项目(3003CB515401)和国际科技合作重点项目(2005FA31110) 作者单位:100730北京,中国医学科学院基础医学研究所、中国协和医科大学基础医学院(△通讯作者)高,其N-端的9,15,20丝氨酸磷酸化水平升高[8]该过程可由各种激酶调节,这些酶调节的p53N端磷酸化可使p53从MDM2解离,达到稳定存在,细胞中p53累积,诱导或者加速细胞凋亡[9]。 在PKCδ诱发的细胞凋亡过程中,p53也是很重要的调节因子。细胞过量表达p53时,可调节细胞周期并引起细胞凋亡,但当利用siRNA技术使该蛋白缺失时PKCδ诱导的细胞凋亡率显著降低[10],但是在单独异位表达p53时,却并不能诱导细胞凋亡,只有与PKCδ共表达,才又再次恢复诱导凋亡能力。研究表明,p53有诸多的磷酸化位点,比如371、376和378等位的丝氨酸,不同位置氨基酸的磷酸化,可引发各不相同的反应。其中, p5315和46位丝氨酸的磷酸化是PKCδ引发细胞凋亡过程中重要的一环。在NO诱使的多巴胺能细胞凋亡过程中,PKCδ因酪氨酸硝化得以激活,进而磷酸化p5315位丝氨酸,磷酸化的p53与MDM2的结合能力减弱,蛋白酶基础降解水平降低,即稳定性增加,蛋白不断积累,进而诱导细胞凋亡[11]。同时PKCδ与p53DINP1结合,将p5346位丝氨酸磷酸化[12],促使形成细胞凋亡诱导蛋白p53AIP1,引起细胞凋亡。 除了与p53蛋白直接反应之外,PKCδ还能通过影响p53的表达来影响细胞的凋亡(同46Ser)。在PKCδ诱导的细胞凋亡中,p53的表达明显升高,而抑制其活性时,p53的基础表达量有了明显的降低[13]。 2磷脂转移酶 磷脂转移酶(PLS)是一种对于细胞线粒体形态、功能以及凋亡应答都具有重要作用的酶。该家族具有四名成员,分别为:位于细胞膜,可转移
【摘要】细胞凋亡是当前生物学领域中研究的最新课题之一。细胞凋亡是个体发育过程中由一系列蛋白调控的细胞主动死亡过程,在保证多细胞生物健康生存的过程中扮演着关键角色,对个体的正常发育具有重要作用。它在多细胞生物的组织分化、器官发育、机体稳态的维持中有着重要意义。其中bcl-2家族、caspase 家族、p53蛋白、survivin蛋白都是重要的凋亡调节因子,在细胞凋亡中相互联系,相互作用,从而调控细胞凋亡.本文探讨了bcl-2家族、caspase家族、p53蛋白、survivin蛋白对细胞凋亡的调控机制。 【关键词】细胞凋亡、 bcl一2家族、caspase家族、p53 蛋白、survivin 蛋白 引言 细胞凋亡是细胞的一种基本生物学现象,在多细胞生物去除不需要的或异常的细胞中起着必要的作用。它在生物体的进化、内环境的稳定以及多个系统的发育中起着重要的作用。细胞凋亡是多蛋白严格控制的过程,随着分子生物学技术的发展对多种细胞凋亡的过程有了较为深入的认识,但是迄今为止凋亡过程确切机制尚不完全清楚。而凋亡过程的紊乱可能与许多疾病的发生有直接或间接的关系。细胞凋亡是一个主动过程,它涉及一系列蛋白的激活、表达以及调控等的作用。其中caspase家族蛋白、Bcl-2家族蛋白和p53蛋白、survivin等在凋亡的信号转导中扮演着重要角色。 一、caspase家族蛋白 1.1 caspase家族蛋白介绍 caspase是半胱氨酸基天冬氨酸一特异性蛋白酶(cystei-nyl aspartate specific proteinase)即半胱氨酸天冬氨酸酶的缩写。Caspase半胱氨酸天冬氨酸特异性蛋白酶(Cysteinyl aspartate specific proteinase,Caspase)家族,也称为ICE/CED-3家族,是美丽线虫(Caenorhabditis elegans)死亡基因CED-3的同源物。这类蛋白酶与细胞凋亡形态学特征变化(如细胞膜空泡形成、核膜破裂、染色质聚集和边聚及DNA断裂等)以及一些生化改变关系密切。它们是一组存在于胞浆中的半胱氨酸蛋白酶,其共同特点是特异性断开天冬氨酸残基后的肽键。到目前为止,在小鼠和人类中,已经发现caspase家族至少有14个成员。细胞中合成的caspase以无活性的酶原状态存在,经活化后方能执行其功能。 1.2 Caspase分类 Caspase分为三大类:凋亡启动因子(apoptotic initiators)、凋亡执行因子(apoptotic executioners)和炎症介导因子(inflammatory mediators),构成了级联放大效应。凋亡启动因子在级联反应的上游,包括Caspase-2、Caspase-8、Caspase-9、Caspase-10等,能在其它蛋白辅助下发生自我活化并识别和激活下游的Caspase。如Caspase-8几乎能激活所有凋亡级联反应下游的Caspase而诱发凋亡。凋亡执行因子在级联反应的下游,包括Caspase-3、Caspase-6和Caspase-7等,作用于其特异性底物并导致细胞凋亡。如Caspase-3,是Caspase家族中的最重要的凋亡执行者之一,是细胞凋亡过程中的主要效应因子。它的活化是凋亡进入不可逆阶段的标志。炎症介导因子包括
胃癌细胞凋亡基因 西安国医肿瘤医院研究人员通过大量的研究分析发现,胃癌细胞凋亡是有相关基因控制的,目前发现的有以下三个基因:bcl-2基因、Bax和Fas/FasL。下面来详细介绍: 1.bcl-2基因 bcl-2基因编码26kD的膜蛋白,是第一个被确认有抑制凋亡作用的基因。bcl-2基因激活、过表达可抑制细胞凋亡,从而使细胞增殖和凋亡不平衡,而且会使具有遗传改变又得不到修复的细胞免于死亡而进入细胞循环,多种遗传成分改变可导致肿瘤的发生。因此,bcl-2在肿瘤发生发展中起着重要的作用。 乳腺癌中,高表达的Bcl-2与乳腺癌细胞凋亡指数呈负相关,而且与有丝分裂指数呈正相关,提示在细胞增殖活跃期,Hcl-2阳性细胞凋亡减少,即Bcl-2过表达影响了细胞凋亡。Nakamum检测了肠型胃癌、胃腺瘤、肠化生及非化生胃黏膜,发现在肠化生中Bcl-2蛋白表达量最高(77.1%),胃腺瘤(37.5%)和肠型胃癌(10.8%)中较低。 因而认为,Bcl-2蛋白的过表达主要是在胃癌的早期阶段起作用,使转化细胞逃避凋亡,以进一步积累其他基因的异常。Lauwers采用单克隆抗体124检测正常、伴有肠化生的萎缩性胃炎及异型增生胃黏膜,发现正常胃黏膜仅在胃小凹与腺体交接处增生的干细胞中有Bcl-2蛋白的微表达,而在肠化生黏膜的过增生区域及胃小凹表面分化不良的细胞中均可检测到Bcl-2,这些分化不良的细胞正是胃癌癌前病变的一个特征。 因此推测,胃黏膜受损后增生加快,导致一些分化不良的细胞出现,这些分化不良的细胞又因Bcl-2蛋白的过表达而逃避凋亡,呈现生长优势,细胞寿命延长,基因变异积累的机会增加,为进一步向恶性细胞转化提供了条件。 2.Bax Bax是第一个被分离到的Bcl-2家族成员之一,与Bcl-2的同源区主要集中在BH1和BH2区。Bax的功能与Bcl-2相对,可促进细胞的凋亡。Bax与Bcl-2在体内的表达呈部位互补形式。Bcl-2倾向于分布在生长细胞、增殖细胞,而Bax倾向于分布在终末分化细胞、退化细胞,在凋亡旺盛的细胞中表达更强。 国外Komatauin报道,在胃黏膜癌变的早期阶段,即已发生Bax的表达异常。
第一节细胞凋亡的生物学意义及其相关基因 对于一个多细胞生物来说,要维持完整性和保持平衡性,凋亡是一个非常重要的生物学过程。多细胞生物的诞生、生长、发育、存活以及死亡,无一不伴随着细胞凋亡过程。 关于细胞增殖能力和寿命是有限的观点。细胞,至少是培养的二倍体细胞,有一定的寿命;它们的增殖能力不是无限的,而是有一定的界限,这就是 Hayflick界限。癌细胞或培养的细胞系是不正常细胞,其染色体数目或形态已经不同于原先的细胞细胞的增殖能力与供体年龄有关。物种寿命与培养细胞寿命之间存在着一定的关系。 一、细胞衰老 二倍体细胞的衰老是由细胞本身决定的。决定细胞衰老的因素在细胞内部,而 不是外部的环境;是细胞核而不是细胞质决定了细胞衰老。在机体内,细胞的衰老和死亡是常见的现象,甚至在个体发育的早期也会发生;衰老动物体内,细胞分裂速度显著减慢,其原因主要是G1期明显延长;衰老个体内的环境因素影响了细胞的增殖和衰老; 二、衰老细胞结构的变化 细胞核的变化: 体外培养的二倍体细胞,细胞核随着细胞分裂次数的增加不断增大;细胞核的核膜内折(invagination)、染色质固缩化。 2. 内质网的变化: 衰老动物内质网成分弥散性地分散于核周胞质中,粗面内质网的总量似乎是减少了。 3.线粒体的变化: 通常细胞中线粒体的数量随龄减少,而其体积则随龄增大;致密体的生成:脂褐质,老年色素等。 4.膜系统的变化: 衰老的细胞,其膜流动性降低、韧性减小。衰老细胞间间隙连接减少;细胞膜内(P面)颗粒的分布也发生变化(减少)三、细胞衰老的分子机理氧化性损伤学说:代谢过程中产生的活性氧基团或分子(ROS---O2-, OH-, H2O2),引发的氧化性损伤的积累,最终导致衰老。
细胞凋亡受基因控制,细胞癌变不受基因控制 1838~1839年,Schleiden和Schwann分别从各自的研究工作中得出一切动物和植物都是由细胞构成的结论,从而创立了细胞学说。1842 年,Vogt在蟾蜍发育过程中观察到正在死亡中的神经细胞。这是人类首次认识在正常生理条件下发生的细胞死亡现象。 1965 年,Lockshin和Williams在他们一系列论文中首次使用“编程性细胞死亡”(progra mmed cell death, PCD)这一术语,用来描述蚕蛾变态期间发生的受时间调节的节间肌退化。这一术语的意思是指存在着一个固有的遗传程序,在受控的时空被激活,导致细胞死亡。1972 用最简单的话说,细胞坏死是细胞的病理性死亡方式;细胞凋亡则是细胞的生理性死亡方式,因此,细胞凋亡常与编程性细胞死亡作为同义词使用。对多细胞生物来说,细胞凋亡负责在胚胎发育过程中清除对机体已经没有用的细胞年,Kerr 等首次使用“细胞凋亡”(apoptosis)一词,用来描述一种与完全由外部因素引起的细胞坏死(n ecrosis)在本质上截然不同的细胞死亡方式的形态特征。apoptosis源自希腊语,意思是凋落或凋谢,如树叶掉落、毛发脱落等,都有选择性地除去一些个别部分而不影响其余部分的特点。(如构成缪勒氏管的细胞对男性是完全没有用的);在成年机体中维持体内的稳定状态,也就是使细胞增殖与细胞消亡两个过程达到平衡;参与机体的防御机制,当受到病毒等病原体侵染时,通过诱导受累细胞的凋亡而减轻其危害。细胞凋亡受阻和细胞凋亡过度都会引起严重的后果,人类的许多严重疾病都与此有关。例如,白细胞凋亡受阻,该死的不死,与白血病有关;免疫活性细胞凋亡过度,不该死的死得太多,与艾滋病有关。 1 细胞凋亡的特征细胞坏死与细胞凋亡的比较见表1。 2 与细胞凋亡有关的主要基因及其产物细胞凋亡是一种受基因调控的主动性编程反应。 人的细胞凋亡机制十分复杂,研究起来十分困难,许多认识来自对秀丽隐杆线虫(Caen orhabditis elegans)的遗传分析。C.elegans营自由生活,长1mm,在20℃培养时的生活周期为3.5天,透明的表皮使每个细胞均可见,最可贵的是细胞数目少且固定,在发育过程中总是产生1090个细胞,其中131个细胞注定要凋亡。用遗传学方法建立了影响C.elegans细胞凋亡的许多突变,从中鉴定了14种与C.elegans细胞凋亡有关的基因,其中,居中心地位的是抑制细胞凋亡的ced-9和促进细胞凋亡的ced-3、ced-4。它们的蛋白产物CED-9、CED-3、CED-4已相继被证明都有人类同源体。此外,哺乳类还发展了一些重要的细胞凋亡机制。 2 .1 Bcl-2家族1985 年,Tsujimoto等将人18q21上涉及滤泡性淋巴瘤的一个癌基因 定名为BCL-2。1988年,Vaux等发现其蛋白产物Bcl-2的功能不是增加细胞的增殖能力而是使细胞凋亡受阻。这是人类第一次认识到细胞凋亡受阻与细胞增殖失控同样能使体内稳态平衡失调而导致肿瘤发生。1994年,Hengartner和Horvitz通过基因结构对比、蛋白结构对比与氨基酸序列对比,确定Bcl-2蛋白是CED-9蛋白的人类同源体。此后陆