
收稿日期:2006210225;修订日期:2007206207
基金项目:国家重点基础研究发展计划(编号:2006C B101805)资助
作者简介:贝锦新(1977—
),男,广东揭西人;博士;主要从事鱼类分子免疫学研究。E 2mail :beijinxin @https://www.doczj.com/doc/762516471.html, 通讯作者:林浩然,E 2mail :lsslhr @https://www.doczj.com/doc/762516471.html, ;T el :020*********
综述
鱼类基因数据库与生物信息学在鱼类基因开发上的应用
贝锦新 张 勇 李文笙 刘晓春 林浩然
(中山大学水生经济动物研究所,广州 510275)
APP LICATION OF GEN OME DATABASES AN D BIOINFOR MATICS
IN EXP LORATION FOR FISH GENES
BEI Jin 2X in ,ZH ANG Y ong ,LI W en 2Sheng ,LIU X iao 2Chun and LIN Hao 2Ran
(Institute o f Aquatic Economic Animals ,Sun Yat 2Sen (Zhongshan )Univer sity ,Guangzhou 510275)
关键词:鱼类基因组;同线性分析;同源比对;基因开发
K ey w ords :Fish gen ome ;C onservation of synteny ;H om ology search ;Data mining
中图分类号:Q78 文献标识码:A 文章编号:100023207(2008)0320387206 近十年来,随着分子生物学技术的发展,各国际组织间的合作研究使得资源基因组计划得以普遍实施。各种动物、植物基因组数据库均在互联网上陆续发布,生物信息量随之迅猛增长,然而如何利用并发掘这些数据蕴涵的宝藏,从中提取解释生命个体生长发育、免疫调节以及病害控制等机理的宝贵信息,已成为人类所面临的巨大挑战。因此,以获取、加工、储存、分配、分析和解读生物信息为手段,综合运用数学、计算机科学和生物学工具的交叉学科———生物信息学[1]由此诞生。也正是依靠生物信息学的辅助,人类基因组计划最终在2003年基本完成,基因组序列数据的拼接和组装也陆续完善。
各种鱼类大约占据了50%的脊椎动物种类,有着长远的进化史
[2]
。因此,利用这些丰富的鱼类资源,研究与系统发育
和进化相关的种间保守或者特异的遗传和分子机制就显得相当重要。但相对于高等脊椎动物而言,鱼类的分子生物学研究比较滞后。在鱼类基因开发过程中,大部分是通过不同鱼类或者是其他高阶元物种的基因序列同源性,设计简并引物进行目的片段扩增;或者通过表达序列标签(Ex pressed se 2
quence tags ,EST )数据库发现并筛选鱼类功能基因。随着鱼类
的基因组全序列测序拼接工作的不断完善,通过生物信息学和分子生物学的方法,新的鱼类功能基因逐渐被发现,这种技术路线采用了反向生物学的原理,即从基因到蛋白质再到功能研究的途径来发现新的生物活性分子[3]。国外科学家已经筛选到一批与发育、生殖及免疫相关的功能基因,而国内对于鱼类新功能基因的开发研究起步较晚,因此,我们应当运用生
物信息学结合分子生物学的手段,发掘鱼类以及其他生物基因组数据库中的关键信息,开发并研究鱼类重要的功能基因,使得我国在这一领域能够有创新性的成果。为此,本文综述了目前鱼类基因组数据库的研究现状,并结合例子介绍利用这些数据库进行基因开发的几种方法。
1 鱼类基因公共数据库资源
111 斑马鱼(Danio rerio ,zebrafish )
斑马鱼隶属鲤形目(Cypriniformes )鲤科,由于个体小、周期产卵且产卵周期短、产卵量高、卵大、体外受精、胚胎透明、体外发育、胚胎早期发育快和易于大量获得样品等独有的特点而成为目前世界公认的模式脊椎动物之一。单倍体斑马鱼基因组由25条染色体组成,含有117×103Mb (1Mb =106个碱基对或bp ),大约为哺乳类基因组大小的二分之一[4]。
2001年2月开始,英国Sanger 研究所开始了斑马鱼全基因组
测序工作;主要通过BAC (Bacterial artificial chrom os ome )、PAC
(P12derived artificial chrom os ome )文库的克隆测序和拼接,以
及基因组鸟枪法测序(Whole genome shotgun sequencing )和组装两种策略。目前,该研究所和国际同行已经测序并拼接了
6653条DNA 片段,包含碱基约116×103Mb (参照:http ://w w w 1sanger 1ac 1uk/Projects/D 2rerio/)。许多不同的服务器都
提供了最新的斑马鱼基因组序列拼接数据,并能进行下载和比对(表1)。同时,以美国华盛顿大学为代表的许多研究所也进行了不同规模的EST 测序工作,这些丰富的斑马鱼EST
第32卷第3期
水生生物学报
V ol.32,N o.32008年5月
ACT A HY DROBI O LOG IC A SI NIC A
May ,2008
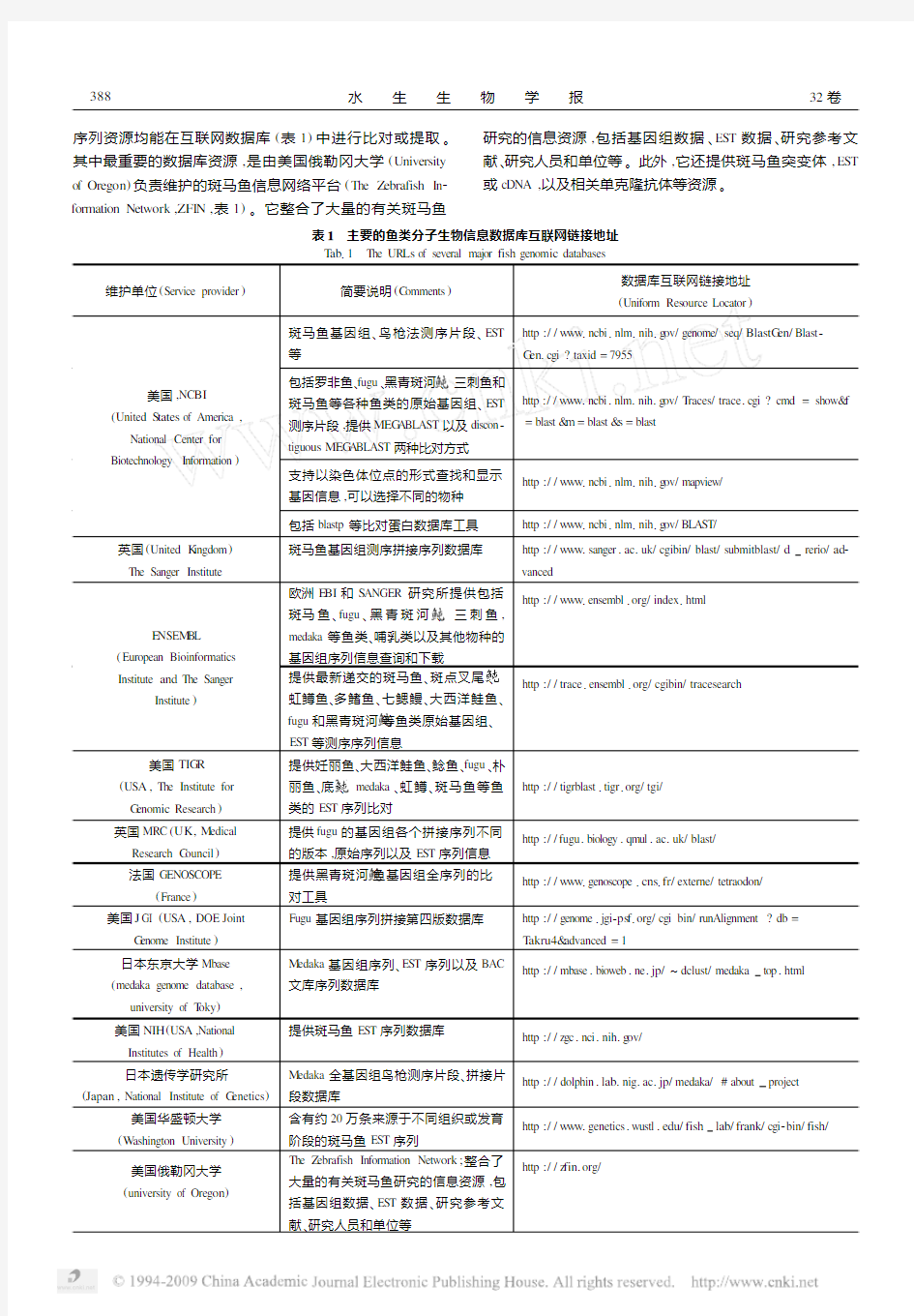
序列资源均能在互联网数据库(表1)中进行比对或提取。其中最重要的数据库资源,是由美国俄勒冈大学(University of Oreg on)负责维护的斑马鱼信息网络平台(The Z ebrafish In2 formation Netw ork,ZFI N,表1)。它整合了大量的有关斑马鱼研究的信息资源,包括基因组数据、EST数据、研究参考文献、研究人员和单位等。此外,它还提供斑马鱼突变体,EST 或cDNA,以及相关单克隆抗体等资源。
表1 主要的鱼类分子生物信息数据库互联网链接地址T ab11 The UR Ls of several major fish genom ic databases
维护单位(Service provider)简要说明(C omments)数据库互联网链接地址(Uniform Res ource Locator)
美国,NC BI (United S tates of America, National Center for Biotechnology In formation)斑马鱼基因组、鸟枪法测序片段、EST
等
http://w w w1ncbi1nlm1nih1g ov/genome/seq/BlastG en/Blast2
G en1cgi?taxid=7955
包括罗非鱼、fugu、黑青斑河 、三刺鱼和
斑马鱼等各种鱼类的原始基因组、EST
测序片段,提供MEG AB LAST以及discon2
tigu ous MEG AB LAST两种比对方式
http://w w w1ncbi1nlm1nih1g ov/T races/trace1cgi?cmd=show&f
=blast&m=blast&s=blast
支持以染色体位点的形式查找和显示
基因信息,可以选择不同的物种
http://w w w1ncbi1nlm1nih1g ov/mapview/
包括blastp等比对蛋白数据库工具http://w w w1ncbi1nlm1nih1g ov/BLAST/
英国(United K ingdom) The Sanger Institute 斑马鱼基因组测序拼接序列数据库http://w w w1sanger1ac1uk/cgibin/blast/subm itblast/d rerio/ad2
vanced
E NSE M BL (European Bioin formatics Institute and The Sanger
Institute)欧洲E BI和S ANGER研究所提供包括
斑马鱼、fugu、黑青斑河 、三刺鱼,
medaka等鱼类、哺乳类以及其他物种的
基因组序列信息查询和下载
http://w w w1ensembl1org/index1htm l
提供最新递交的斑马鱼、斑点叉尾 、
虹鳟鱼、多鳍鱼、七鳃鳗、大西洋鲑鱼、
fugu和黑青斑河 等鱼类原始基因组、
EST等测序序列信息
http://trace1ensembl1org/cgibin/tracesearch
美国TIG R (US A,The Institute for G enom ic Research)提供妊丽鱼、大西洋鲑鱼、鲶鱼、fugu、朴
丽鱼、底 、medaka、虹鳟、斑马鱼等鱼
类的EST序列比对
http://tigrblast1tigr1org/tgi/
英国MRC(UK,M edical Research C ouncil)提供fugu的基因组各个拼接序列不同
的版本,原始序列以及EST序列信息
http://fugu1biology1qmul1ac1uk/blast/
法国GE NOSCOPE
(France)提供黑青斑河 鱼基因组全序列的比
对工具
http://w w w1genoscope1cns1fr/externe/tetraodon/
美国J GI(US A,DOE Joint
G enome Institute)Fugu基因组序列拼接第四版数据库http://genome1jgi2ps f1org/cgi bin/runAlignment?db=
T akru4&advanced=1
日本东京大学Mbase (medaka genome database, university of T oky)M edaka基因组序列、EST序列以及BAC
文库序列数据库
http://mbase1bioweb1ne1jp/~dclust/medaka top1htm l
美国NIH(US A,National Institutes of Health)提供斑马鱼EST序列数据库
http://zgc1nci1nih1g ov/
日本遗传学研究所(Japan,National Institute of G enetics)M edaka全基因组鸟枪测序片段、拼接片
段数据库
http://dolphin1lab1nig1ac1jp/medaka/#about project
美国华盛顿大学(W ashington University)含有约20万条来源于不同组织或发育
阶段的斑马鱼EST序列
http://w w w1genetics1wustl1edu/fish lab/frank/cgi2bin/fish/
美国俄勒冈大学(university of Oreg on)The Z ebrafish In formation Netw ork;整合了
大量的有关斑马鱼研究的信息资源,包
括基因组数据、EST数据、研究参考文
献、研究人员和单位等
http://z fin1org/
388
水 生 生 物 学 报32卷
112 青 (Oryzias latipes,Medaka)
青 是一种淡水颌针目(Beloniformes)鱼类;体型小、生长周期短、繁殖力强、卵透明,对于水中溶氧及温度的变化适应能力较强;可作为环境、癌症以及发育等研究领域的一种实验对象[5]。青 的基因组由24对染色体组成,含有碱基约(0165-1)×103Mb;其大小相当于人类基因组的三分之一、小于斑马鱼的二分之一[6]。日本科学家成功地获得许多自然的或者人为诱导的(E thylnitros ourea,E NU)突变株;最近,他们成功地获得了透明青 突变株[7]。2002年开始,日本研究团体共同实施了青 基因组计划,包括进行大规模EST测序、BAC文库构建测序、全基因组鸟枪法测序以及基因遗传图谱制作等等。目前,来源于不同青 cDNA文库的将近10万条EST序列收录于日本东京大学管理的公共数据库(Mbase,表1)中,为研究者提供比对分析工具。大约90%的基因组序列已经测定,而拼接工作也在不断完善;日本遗传学研究所(National Institute of G enetics)提供了有关的基因组序列数据库(表1)。青 本身具有的特征以及目前取得的研究成果,将逐渐使它成为一种新的模式脊椎动物。
113 红鳍东方 (Takifugu rubripes,fugu)
红鳍东方 ,俗称河 鱼,为 形目(T etraodontiformes)海水鱼类,但对盐度变化具有较强地适应能力。河 体型较大,繁殖周期比较长,性成熟大约需要3年,卵不透明[8]。其基因组大小为400Mb,不到人类基因组的七分之一[9];虽然它的基因组比较小,但作为一种脊椎动物,它具有与哺乳类许多相同的生物学特征,因此也具有许多相似的基因[10]。Brenner,et al1[9]研究发现河 鱼的基因结构比较紧凑(C om2 pact),也就是部分基因的内含子序列较短或少,而且基因之间间隔较短;并首次提出红鳍东方 可以作为脊椎动物基因研究的模式生物[9]。E lgar,et al1[11]以及Aparicio,et al1[12]陆续报道了河 鱼的基因组测序和相关的拼接研究工作。在Aparicio,et al1的研究基础[12]上,到2004年8月,J GI(The DOE Joint G enome Institute)以及其他研究组利用质粒、粘粒和BAC序列,完成了大于95%覆盖率的基因组序列拼接工作。他们最新的数据库(表1)提供了河 鱼基因组的7213条组装序列,含有大约393Mb。伦敦大学玛丽皇后学院(Queen Mary,University of London)开发了利用河 鱼基因组序列信息以及大量的EST序列信息进行目的基因比对的生物信息学工具(表1)。
114 黑青斑河 (T etraodon nigroviridis,S potted2green pu ffer fis) 2004年,Jaillon,et al1[13]报道了黑青斑河 的基因组序列草图。黑青斑河 是 形目淡水鱼类,体型小,可在小型水族箱中饲养。它与红鳍东方 的亲缘关系很近,由18—30百万年前的共同祖先进化而来[13];它的基因组大小为350Mb,是已知基因组最小的脊椎动物。目前有39条超长拼接序列(Ultracontigs)已经定位到该种的21条染色体上,覆盖了基因组的6412%(参照:http://w w w1gen oscope1cns1fr/externe/te 2 traod on/)。此外,Jaillon,et al1[13]也提供了大量的EST序列信息,可以通过GE NOSC OPE、E NSE M BL或者NC BI数据库工具进行比对和提取(表1)。
115 其他种类
此外,以美国、英国、日本等国为主的基因组中心分别开展了各种鱼类的基因组和EST测序工作。目前,包括刺鱼形目的三刺鱼(Gasterosteus aculeatus,Three2spine stickleback)、鲶形目的斑点叉尾 (Ictalurus punctatus)、鲑形目虹鳟鱼(On2 corhynchus mykiss,Rainbown trout)和大西洋鲑鱼(Salmo salar)、多鳍鱼目多鳍鱼(Polypterus bichir)、鲈形目的妊丽鱼(Astatoti2 lapia burtoni)和朴丽鱼(Haplochromis sp1red tail sheller)、 形目的底 (Fundulus heteroclitus,killifish)以及圆口纲七鳃鳗(Petromyzon marinus)在内的许多鱼类基因序列信息,都能在TGI、E NSE M BL或者NC BI等公共数据库服务器中得以查询提取(表1)。随着这些种类基因序列信息的不断完善,将为比较基因组学和比较生物学提供极其重要的平台。
2 利用基因数据库开发鱼类新基因
一般而言,鱼类基因的克隆以及功能研究要比高等哺乳类的相关研究滞后许多。近几年,得益于鱼类基因组数据库的不断更新完善,从事鱼类研究工作的科学家开始重视这些数据库的功能,并利用它们开发新型的鱼类基因。根据目前的报道,利用基因数据库开发新基因的方法大致有两种,包括基因编码蛋白序列相似性的同源比对法和基因组片段同线性进化现象的同线性分析法
。以下分别列举一个例子,介绍如何运用这两种方法。
211 同源比对
物种间基因的比较表明,生物在进化过程中,不同种属之间的同一基因的编码序列以及基因结构具有保守性,表现为一定的同源性。根据这个特点,可以利用已知物种基因的编码氨基酸序列在鱼类基因数据库中搜索候选基因,然后运用生物信息学结合分子生物学的方法,筛选并克隆鱼类的同源目的基因(H om ologue)。主要过程(图1),以最近K aslin,et al1[14]报道的鱼类开胃素(Orexin)基因的克隆为例子。首先,将同源的人orexin基因氨基酸序列输入斑马鱼基因数
图1 同源比对方法开发鱼类新基因的策略
Fig.1 The strategy of hom ology searching for fish genes
in genom ic database
3期贝锦新等:鱼类基因数据库与生物信息学在鱼类基因开发上的应用389
据库(S A NG ER ,表1)比对框,选择T B LAST N 程序,E 值(E x pect )=
1进行查询(图2)。图3结果显示有一个含有候选基因片段的
拼接序列,选择并下载含有该候选基因的邻接序列(C ontig )。然后,利用G E NS C A N 软件[15](http ://genes.m https://www.doczj.com/doc/762516471.html,/G E NS C A N.htm l )对该序列进行基因预测并将预测基因输入NC BI G en Bank 数据库进行blastp 比对;其中,得到具有一段orexin 蛋白结构(图4)的预测基因。接着,以这段预测的cD N A 为基础,设计特异引物进
行全长cD N A 克隆,或者采取3′和5′2R ACE (Rapid am plification of
cD N A ends )的方法克隆目的基因。最后,通过蛋白的氨基酸序
列分析、同源基因的基因结构和表达模式相似性或者结合功能分析去验证该目的基因[14]。经比较,预测的斑马鱼orexin 基因包含所有编码成熟肽的cD N A 序列,说明这种方法的准确性比较高。目前,有许多新的鱼类基因是通过这种方法发现的,如
S OCS 23[16],II 型细胞因子及受体[17]
等。
图2 斑马鱼基因组数据库查询界面(互联网链接见表1)
Fig.2 The query inter face of zebrafish genome database (See T ab.1for UR L
)
图3 斑马鱼基因组数据库查询部分结果
Fig.3 Results for the queried sequence.The underlined zG 210G 13is the name and link to a contig zC210G 13为候选拼接序列名称,单击进入下载页面,P (N )值一般情况下取最低值。Frame 栏表示目
的基因编码方向及起始位点,-2即表示基因方向与拼接序列方向相反,在2号位点起始编码
zC210G 13is the candidate contig ,and could be downloaded via the link.The lowest P (N )value stands for highest probability.The -2in Frame column indicated that the orientation of gene coding was reverse to the candidate contig
sequence and the codon started at the second
position
图4 预测基因BLASTP 比对NC BI 蛋白数据库结果
Fig.4 Results for the predicted protein using B LASTP and protein database in NC BI
上方数字标尺为预测基因编码蛋白长度,下方为orexin 蛋白结构及相对应区域 Ruler above showed the length of predicted protein and below showed the known protein m otif
390 水 生 生 物 学 报32卷
212 同线性分析
同源比对的方法适用于保守程度比较高的基因,但对于一些高变异的基因,如大部分的白细胞介素和一些激素类[18,19],就找不到候选片段。
比较基因组学研究发现,处于不同进化地位的生物,他们当中的许多单个基因不但在编码序列、调控序列以及基因结构上具有保守性,而且基因间的位置关系也具有一定的保守性。也就是说,一些连锁的基因在进化历程中,总是组成
相对稳定的片段,在这些片段中的基因线性排列顺序以及结构高度相似,这种现象被称为“保守同线性(C onserved synte2 ny)”[10,13,20]。物种间分化时间越短,同线性基因越多,区域越长,同线性程度越高。例如人和鼠之间含有417个大于300kb的同线性区域[20],而人和河 鱼享有900个含有2—55个基因的同线性区域[13]。生物体基因组进化的这种现象,为克隆某些高变异基因如大多数的细胞因子提供了一种途径。最近几年,这种方法被广泛用于开发鱼类的新基因,如白细胞介素22、210、215、221,leptin等[19,21—23]。具体的策略(图5),以河 鱼白细胞介素(I L)215的基因克隆[21]为例子。首先,通过NC BI Map viewer(表1)确定人类四号染色体上的I L215基因以及其两侧基因的顺序和方向(
图6);同样,也能确定鸟类的同源基因,发现I L215基因区域存在同线性(图6)。然后,我们通过同源比对方法,分别在河 鱼数据库中找到除了I L215外的其他几个同源基因,而且发现它们呈同线性分布在几个相邻的拼接序列中。第三步,下载河 鱼 Z NF330和I NPP4B基因之间的DNA序列并运用GE NSC AN进行基因预测;经BLASTP比对NC BI蛋白数据库,结果显示,该序列编码一个与I L215同源的基因。第四步,根据预测的cDNA设计特异性引物,分别进行3′和5′2RACE,得到编码I L2 15的全长cDNA。最后,根据氨基酸序列相似性、类似的蛋白高级结构、相同的基因结构以及相似的基因表达模式等特点,我们认为该候选基因为人类I L215的直向同源基因[21]。
图5 同线性分析法开发鱼类新基因的策略
Fig.5 The strategy of conserved synteny analysis of
fish genes in genom ic database
图6 I L215及旁侧基因的同线性
Fig.6 The conservation of synteny of I L215loci
方框内为各个基因缩写,箭头表示基因在基因组片段中的方向,虚线表示同源基因(摘自文献[21]) The genes were boxed,the orientation was shown by arrow,and the hom ologues were linked by dashes(Reference[21])
3 总结与展望
在进行鱼类新基因开发时,首先可以通过查询各种鱼类EST数据库,筛选同源的基因序列[1]。如果这种方法未能得到充分的数据,可以结合本文中的两种方法,找到模式鱼类的同源目标基因。对于那些没有充分的EST或者基因组序
3期贝锦新等:鱼类基因数据库与生物信息学在鱼类基因开发上的应用391
列信息的实验鱼类,可以通过对以上几种模式鱼类同源基因的比对,确定这些基因是否存在保守的功能区域,据此降低引物或者探针设计的简并程度,提高特异性,然后进行PCR 或者文库筛选克隆那些鱼类的同源基因。如果同源性不高以至于不能设计简并引物,那么可以利用基因组片段的同线性现象,选用该同源基因旁侧(最好是两侧)保守基因的编码序列:设计PCR引物,尝试反向PCR的方法对基因组进行步移[24];或者标记作探针,去筛选目的鱼类基因组文库中对应于该区域的基因组片段,将该片段测序,再进行基因预测,找到该区域的目的基因。
另外,在查找某些基因的时候,如果出现两个以上的候选序列,可以根据同源蛋白功能特征序列对每个预测基因进行初步筛选。一个同源基因,也可能在鱼类出现两个或以上拷贝,因为目前的研究说明了鱼类分支在进化过程中发生了一次全基因组复制(Whole genome duplication)[13]。例如,我们还发现一种鱼类特有的I L215类似基因(I L215L),但在基因表达模式上与I L215不同,具体功能上的差异,仍有待于进一步的功能研究[21]。
总之,这里介绍的方法,为鱼类的基因开发提供了一种途径,但对于一些特殊的高变异基因(如白细胞介素23,24, 215等),通过同源性比对的方法未能发现;而且同线性分析也同样未能开发这些基因,这可能受限于基因组序列信息的不完整性,因此,有待鱼类基因组数据库的不断更新和完善。随着更多物种的EST和基因组测序计划的实施,鱼类分子生物学数据库将不断丰富起来,我们将可以更有效地利用生物信息学工具、比较基因组学等研究方法去开发与鱼类生殖、生长、免疫调控相关的功能基因。
参考文献:
[1] Zhao G P.Bioin formatics[M].Beijing:Academ ic Press.2002,3—
12[赵国屏.生物信息学.北京:科学出版社.2002,3—12] [2] Nels on J S.Fishes of the w orld[M].New Y ork:John W iley and
S ons Inc.1994,2
[3] Liu H T.C loning and the strategy of new cytokines[J].International
Journal o f Immunology,1995,5:256—261[刘红涛.近年新细胞
因子的克隆及其技术路线.国外医学(免疫学分册),1995,
5:256—261]
[4] P ostlethwait J H.Ev olution of the Z ebrafish G enome[A].In:G ong
Z,K orzh V(Eds.),Fish Development and G enetics[C].W orld
Scientific,2004,581—611
[5] M itani H,Shima A,Naruse K,et al.M edaka genome mapping to
functional genom ics[A].In:G ong Z,K orzh V(Eds.),Fish Devel2
opment and G enetics[C].W orld Scientific,2004,612—636
[6] W ittbrodt J,Shima A,Schartl M.M edaka2a m odel organism from the
far East[J].Nat Rev G enet,2002,3:53—64
[7] W akamatsu Y,Pristyazhnyuk S,K inoshita M,et al.The see2through
medaka:a fish m odel that is transparent throughout life[J].Proc
Natl Acad Sci U S A,2001,98:10046—10050
[8] Sun Z Z.Biological characters and artificial breeding of pu ffer fish[J].
Shandong F isheries,2002,19:44—46[孙中之.红鳍东方 的生
物学特性及人工育苗技术.齐鲁渔业,2002,19:44—46]
[9] Brenner S,E lgar G,Sand ford R,et al.Characterization of the
pu ffer fish(Fugu)genome as a com pact m odel vertebrate genome[J].
Nature,1993,366:265—268
[10] Liu S M,Y ang L.Pu ffer fish2An ideal m odel for genetic studies of ver2
tebrate[J].Int J G enet,1998,21:1—4[刘山鸣,杨立.脊椎动
物基因研究的理想模式生物2河豚鱼.国外医学遗传学分册,
1998,21:1—4]
[11] E lgar G,C lark M S,M eek S,et al.G eneration and analysis of25Mb
of genom ic DNA from the Pu ffer fish Fugu rubripes by sequence scan2
ning[J].G enome Research,1999,9:960—971
[12] Aparicio S,Chapman J,S tupka E,et al.Whole2G enome Shotgun
Assembly and Analysis of the G enome of Fugu rubripes[J].Science,
2002,297:1301—1310
[13] Jaillon O,Aury J M,Brunet F,et al.G enome duplication in the
teleost fish Tetraodon nigroviridis reveals the early vertebrate proto2
kary otype[J].Nature,2004,431:946—957
[14] K aslin J,Nystedt J M,Ostergard M,et al.The orexin/hypocretin
system in zebrafish is connected to the am inergic and cholinergic sys2
tems[J].J Neuro sci,2004,24:2678—2689
[15] Burge C,K arlin S.Prediction of com plete gene structures in human
genom ic DNA[J].J Mol Biol,1997,268:78—94
[16] Jin H J,Shao J Z,X iang L X.Identification and characterization of
suppress or of cytokine signaling3(S OCS23)hom ologues in teleost fish
[J].Mol Immunol,2007,44:1042—1051
[17] Lutfalla G,Crollius H R,S tange2Thomann N,et al.C om parative ge2
nom ic analysis reveals independent expansion of a lineage2specific gene
fam ily in vertebrates:the class II cytokine receptors and their ligands
in mammals and fish[J].BMC G enomics,2003,4:29
[18] Harris on G,W edlock D.M arsupial cytokines:structure,function and
ev olution[J].Dev Comp Immunol,2000,24:473—484
[19] K urokawa T,Uji S,Suzuki T.Identification of cDNA coding for a ho2
m ologue to mammalian leptin from pu ffer fish,Takifugu rubripes[J].
P eptides,2005,26:745—750
[20] Bourque G,Pevzner P A,T esler G.Reconstructing the G enom ic Ar2
chitecture of Ancestral M ammals:Less ons From Human,M ouse,and
Rat G enomes[J].G enome Res,2004,14:507—516
[21] Bei J X,Suetake H,Araki K,et al.T w o interleukin(I L)215hom o2
logues in fish from tw o distinct origins[J].Mol Immunol,2006,43:
860—869
[22] Bird S,Z ou J,K ono T,et al.Characterization and expression analy2
sis of interleukin2(I L22)and I L221hom ologues in the Japanese
pu ffer fish,Fugu rubripes,following their discovery by synteny[J].
Immunogenetics,2004,56:909—923
[23] Z ou J,C lark M S,Secombes C J.Characterisation,expression and
prom oter analysis of an interleukin10hom ologue in the pu ffer fish,
Fugu rubripes[J].Immunogenetics,2003,55:325—335
[24] Jong A Y,T ang A,Liu D P,et al.Inverse PCR:G enom ic DNA
C loning[A].In:Chen B Y,Janes H W(Eds.),PCR C loning Pro2
tocols(2nd)[C].New Jersey:Humana Press.2002,301—307
392
水 生 生 物 学 报32卷
罗非鱼鱼苗的饲料配方和相关的加工技术要求 一:饲料配方设计的思路 1.明确目标:成本最小,效益最大化 3.罗非鱼鱼苗生长阶段的消化生理特点 幼鱼的食物则以动物性食料(浮游动物)占的比重较大。当鱼长到5厘米时,就逐步向成体的食性过渡。对蛋白质的含量要求较高。 4.饲料原料的营养成分及其特性,相关成本如下: 原料粗蛋白质粗脂肪粗纤维粗灰分总磷价格(元/kg) 鱼粉62.6 9.2 1.5 20.8 2.9 5.5 豆粕44 2 6 6.1 0.62 2.2 花生饼44.7 7.2 5.9 5.1 0.53 3 棉子粕42.6 2.9 11.9 6.7 11.7 2.8 全脂棉籽21.2 19.1 23.4 3.9 6.3 11.5 次粉15.4 2.2 1.5 1.5 0.48 2.1 小麦麸15.7 3.9 8.9 4.9 0.92 0.73 干草15 3.1 23.3 7.8 2.8 6.5 磷酸二氢 钙0 0 0 0 23 2.5 预混料 1 1 3 0 2 8 鱼粉,豆粕,棉子粕等蛋白质,粗纤维含量较高,其饲料特性是饲料柔软,结构疏松,易于消化吸收。 二:饲料配方设计的原则:科学性,实用性,经济性,安全性。 三:罗非鱼鱼苗的饲料配方的制作方法 罗非鱼鱼苗饲料配方 原料比例% 粗蛋白 质 粗脂 肪 粗纤 维粗灰分 总 磷价格(元/kg) 鱼粉24 62.6 9.2 1.5 20.8 2.9 5.5 豆粕25 44 2 6 6.1 0.6 2.2 花生饼20 44.7 7.2 5.9 5.1 0.5 3 棉子粕 1 42.6 2.9 11.9 6.7 12 2.8 全脂棉籽 1 21.2 19.1 23.4 3.9 6.3 11.5 次粉15 15.4 2.2 1.5 1.5 0.5 2.1 小麦麸12.5 15.7 3.9 8.9 4.9 0.9 0.73 干草0.1 15 3.1 23.3 7.8 2.8 6.5 磷酸二氢钙 1 0 0 0 0 23 2.5 预混料0.4 1 1 3 0 2 8 合计100 理论值39.8935 5.193 4.766 8.484 1.6
罗非鱼池塘养殖技术 池塘养殖罗非鱼,成本低,花劳力少,饲养管理简便。在长江流域,南方一般有5~6个月的生长期。只要充分利用生长期,合理放养,加强施肥、投饲,当年鱼种完全可以养成商品鱼,而且可以获得高产。 罗非鱼种苗投放篇 一、池塘条件:罗非鱼的池塘养殖,对池塘条件没有特殊要求,一般养殖家鱼的池塘或农村中的小水塘、沟渠都可以。池塘应选择在水源充足,注排水方便的地方。面积不要超过10亩。因为池塘过大,水质不易肥沃,而且不易捕捞。水深一般1.5~2米。放养鱼种前,池塘要进行清整、消毒和肥水。 二、鱼种放养:鱼种规格要尽量整齐,体长要在3厘米以上的吉富快大罗非苗,体质健壮,无伤无病。鱼种放养时间,要根据当地的气温和水温而定,只要水温稳定在18℃以上,就可以放养。在季节上以提早放养为好,这样可以延长罗非鱼的生长期,提高商品鱼规格和产量。 三、养殖方式:池塘养殖罗非鱼,可以单养,也可以混养,都能获得高产和效益,但以混养效果更好。 1、单养:池塘单养罗非鱼,方法比较简便。养殖措施,可采取一次放养,分次捕捞,捕大留小的方法。放养密度,一般水深1~1.5米深的池塘,每亩放养早繁鱼种3000~4000尾或越冬鱼种2500尾左右,饲养5个多月,每尾可达250克,一般亩产400~500公斤,高的可达700公斤以上。 2、混养:罗非鱼和“四大家鱼”及鲤鱼等品种进行同池混养,可以提高饲料、肥料的利用率,改善水质,并能发挥与其它鱼类的互利作用,而达到促进生长,提高效益。混养方式有两种:
一种是以罗非鱼为主,混养其它鱼类。放养密度,一般每亩放养罗非鱼早繁鱼种2000~2500尾,或越冬鱼种1500~2000尾。其他鱼:鲢(规格250克/尾)250尾,鳙(仙格250克/尾)30-40尾,草鱼(规各500克/尾)50尾,鲤鱼(规格13厘米/尾)10尾。亩产塘鱼可达到600~800公斤,其中罗非鱼占70%~80%。 另一种以家鱼为主,混养罗非鱼。利用水质较肥的池塘,在不降低主养鱼放养密度情况下,放养一定数量的罗非鱼。放养数量随各地养殖方法不一样而不同。一般在亩产750公斤的高产鱼池中,每亩可混养罗非鱼越冬鱼种400~600尾,或混养早繁鱼种800~1200尾,一般可亩产罗非鱼150公斤以上。 罗非鱼饲养管理篇 1、施肥:饲养罗非鱼不论是单养还是混养,都要求水质肥沃。施肥主要是培养水中的浮游生物供罗非鱼摄食,同时肥料的沉底残渣也可以直接作为罗非鱼的饵料。一般施肥量为每周每亩施粪肥200~300公斤或绿肥300公斤左右。施肥要掌握少而勤的原则。施肥的次数和多少,要根据水温、天气、水色来确定。水温较低,施肥量可多些,次数少些;水温较高,施肥量要少,次数多些。阴雨、闷热天气,少施或不施,天睛适当多施。水色为油绿色或茶褐色,可以少施或不施肥;水色清淡的要多施。 2、投饲:池塘施肥培育天然饵料还不能满足罗非鱼的生长需要,必须投喂足够的人工饲料才能获得高产。一般每天上午9时,下午2时各投喂喂料1次,饲料要新鲜,霉烂变质的不能喂。豆饼、米糠等要浸泡后再喂。饲料要投放在固定的食场内。每天投 喂量要根据池塘水温、天气和水质及鱼吃食情况而掌握。一般每次投饲后在1~2小时内吃完为宜。晴天,水温高可适当多投喂;阴雨天或水温低,少喂;天气闷热或雷阵雨前后应停止投喂。一般肥水可正常投喂,水质淡要多喂,水肥色浓要少喂。 3、日常管理:每天早、晚要巡塘,观察鱼的吃食情况和水质变化,以便决定投饲和施肥的数量。发现池鱼浮头严重,要及时加注新水或增氧改善水质。 罗非鱼病害防治
罗非鱼类立克次体病的主要症状和流行特点 我国台湾地区曾在1992年暴发罗非鱼类立克次体病,平均死亡率达到30%,严重发病的达到90%。近几年,我国云南省南部、中部地区养殖的罗非鱼也发生类立克次体病,笔者将云南省罗非鱼类立克次体病的主要发病症状和流行情况做一总结,供相关人员参考。 立克次体为一类严格细胞内寄生的原核细胞型微生物,鱼类立克次体病是欧美国家养殖鲑鳟鱼类的重要疾病,造成巨大经济损失。我国台湾省报道1992年罗非鱼类立克次体病暴发流行,平均死亡率达到30%,严重发病的多个渔场,死亡率甚至达到90%。 近几年,我国云南省南部、中部地区养殖的罗非鱼也发生一种相似的疾病,经过分子生物学检测、组织病理切片观察和发病症状、流行特征观察分析,初步诊断也是类立克次体病。本文把云南省罗非鱼类立克次体病的主要发病症状和流行情况做一总结,供相关人员参考。 一、罗非鱼类立克次体病的临床症状 1、水体中症状 发病鱼上浮、靠边,中午和下午上到水面下停滞,俗称“停尸”。有的发病鱼死亡前常常打转,这与立克次体能感染脑部组织有关。 2、体表症状 多数发病鱼体表没有明显异常症状,有的发病鱼在鳍基部、头部鳃盖等部位有白色结节,有的病鱼鳍基、鳃盖等部位溃疡。立克次体病是慢性病,发病后长期不进食,发病时间较久的鱼体质明显瘦弱。 3、解剖症状 典型症状是在肾脏、脾脏形成大小不等的白色结节,肝脏上白色结节少见,有的病鱼在心脏、肠道也形成白色结节;脾脏肿大,表面形成绿色泡状结构;有的病鱼脑部发红、充血;多数发病鱼有腹水;发病时间较长以后内脏粘连。 4、罗非鱼类立克次体病与诺卡氏菌病症状上的区别 目前未见罗非鱼诺卡氏菌病的报道,罗非鱼类立克次体病与其它鱼类的诺卡氏菌病有很大区别: (1)罗非鱼类立克次体病有打转、身体僵硬、异常游泳等神经症状,与立克次体造成脑部感染有关。而诺卡氏菌一般不能感染脑部,所以一般没有神经症状。 (2)罗非鱼类立克次体病体表有白色结节,肌肉没有白色结节。诺卡氏菌病体表一般不形成白色结节,但可以在肌肉形成白点结节。
附件 第四届全国水产原种和良种审定委员会 第五次会议审定通过品种简介 一、品种登记说明 全国水产原种和良种审定委员会审定通过的品种登记号说明如下: (一)“G”为“国”的第一个拼音字母,“S”为“审”的第一个拼音字母,以示国家审定通过的品种。 (二)“01”、“02”、“03”、“04”分别表示选育种、杂交种、引进种和其他类品种。 (三)“001”、“002”、……为品种顺序号。 (四)“2012”为审定通过的年份。 如:“GS-01-001-2012”为凡纳滨对虾“桂海1号”的品种登记号,表示2012年国家审定通过的排序1号的选育品种。 二、品种简介 (一)品种名称:凡纳滨对虾“桂海1号” 品种登记号:GS-01-001-2012 亲本来源:美国凡纳滨对虾选育群体 选育单位:广西壮族自治区水产研究所 品种简介:该品种是以2006年从美国引进的凡纳滨对
虾选育群体为基础群,采用家系选育技术,以生长速度和养殖成活率为选育指标,年建立家系60个,选留家系12个,每家系按5%留种率选留600尾后代,经连续5代选育而成。 在5万尾/亩的放养密度下,与从美国进口种虾生产的第一代虾苗相比,该品种生长速度快,单造亩产量可提高13.97%;成活率高,单造养殖成活率可达80.88%,提高11.32%以上;85日龄后展现出明显生长优势,130日龄平均体重提高15%以上。 适宜在我国各地人工可控的海水、咸淡水水体中养殖。 (二)品种名称:三疣梭子蟹“黄选1号” 品种登记号:GS-01-002-2012 亲本来源:野生三疣梭子蟹 选育单位:中国水产科学研究院黄海水产研究所,昌邑市海丰水产养殖有限责任公司 品种简介:该品种是以2005年收集的莱州湾、鸭绿江口、海州湾和舟山4个野生三疣梭子蟹群体构建基础群,以生长速度为选育指标,经连续5代群体选育而成。 与未经选育的三疣梭子蟹相比,在相同条件下进行养殖,收获时该品种平均体重可提高20.12%,成活率可提高30.00%,且全甲宽变异系数小于5%,规格整齐。 适宜在浙江及以北沿海人工可控的海水水体中养殖。 (三)品种名称:“三海”海带
目前钓罗非鱼使用的冷冻饵分类:腥味、肝味两大类。DMPT TMAO甜菜碱丙氨酸 一、开冷冻饵步骤→钓腥味 ①、将腥味冷冻饵1包自然解冻; ②、解冻完后,分成4块倒入饵料盆内,横向捏开分散; ③、再倒入1/3量的赤尾青(小袋装)与冷冻饵一起拌均即可。 二、香料: ①、3勺量(陶瓷汤匙)丸九料(小袋装香草味红鲫)1份; ②、1勺量丸九料3#鲫(草莓香型);一起另装盆混合拌均,再加1汤匙水,然后 迅速搅拌,任其吸收3-5分钟待用。 三、香精: ①、香虎量(黄豆大小)1份; ②、蛋奶量(小手指1节大小,台湾牌深蛋黄色)1份; ③、牛奶香量是蛋奶量的一半; ④、猪母奶量(比小黄豆小,不宜多加); ⑤、鸡精(家乐牌或太太乐牌5-10粒); ⑥、将以上香精一起倒入溶器内用水溶解(建议温水为佳),然后倒入【配料二 :香料】,将它们一起拌均(料的湿度大点为宜); ⑦、待丸九料充分吸收香味后,再将其倒入【配料一:冷冻饵】内一起拌均,充 分使冻饵与香料相溶; ⑧、然后掺入1/3量的好小子虾粉以及1/20量的丸九牌虾粉(小红袋装),将它们 一起搅拌均匀。 这样就调成您所需的罗非香型钓饵,最后再将其急速冷冻,待第2天使用或现
场开料使用。 (注:制成饵料后,钓鱼时先捏起一小块饵料,抛入盛水盆内观察饵料雾化 状态是否是您所需的时间效果,要想增加粘度,上饵料时多揉捏些时间便可,要增加雾化相反。) 四、开罗非饵腥香型饵料,要根据天气变化而定香味。 ①、热天(气温28度-38度),适合使用草莓香与草香混合; ②、冷天或雨天(气温18度-28度),适合使用凤梨香型或奶香型。钓鱼时要及时变化,找出适合鱼儿的香型。吃腥味时,另把虾粉一包或半包倒入罗非香饵内搅均即可。 例如: ①、好小子腥味1包自然解开,开料顺序同上; ②、香料:丸九红鲫3汤匙量,不加丸3#; ③、香精:巧克力香精和牛奶香精1拇指量大,香虎、鸡精、猪母乳量及顺序同上,这样配法就制成腥香型巧克力罗非钓饵,效果佳。 五、开冷冻饵肝味步骤→钓肝味,开料方法及顺序与腥味开料方法及顺序一样;不过,香味份量加大方可。如: ①、冷冻饵肝味1包(建议使用“五福”肝味或“鱼神牌肝味); ②、香料:6汤匙丸九牌红鲫、1勺半(陶瓷汤匙)丸九3#鲫; ③、赤尾青1/2袋量(小袋装); ④、香虎量(比黄豆大),蛋奶、牛奶量是腥料的一倍,猪母奶量、鸡精量同【 开腥味料】用量一样。这样就制成肝香型罗非型钓饵。 六、肝腥混合香型钓饵:
(一)复合添加剂配方? 1?杂食性鱼类复合添加剂配方 杂食性鱼复合添加剂是根据杂食性鱼类生物学特点设计而成,一般分为矿物质添加剂和维生素添加剂两种。? (1)杂食性鱼类复合添加剂配方? 鲤鱼罗非鱼: 矿物质或微量元素(mg):每克预混料含铁50,铜3,钴0.01,锰20,锌30,碘0.1,硒0.01,饲料中添加量为0.1 微生素混合物组成(mg):每克预混料含A为1000IU,E为10IU,K2为2,B1为4,B2为4,泛酸10,尼克酸20,吡哆醇4,叶酸1,C为40,B12为0.004,生物素0.02,氯化胆碱90,添加量为0.6% 鲤鱼: 矿物质或微量元素(mg):每克预混料含铁100,锰50,锌50,铜10,钴0.05,碘0.1,添加量为0.1% 微生素混合物组成(mg):每克预混料含A为5000IU,D600IU,B1为10,B2为20,泛酸30,尼克酸50,抗坏血酸200,添加量为0.1% (2)肉食性鱼类复合添加剂配方? 鲶鱼: 矿物质或微量元素(mg):每克预混料含锰115,锌88,铁44,铜10,碘0.3,添加量为0.1% 微生素混合物组成(mg):每克预混料含A5500IU,D1000IU,E为50,K为10,B1为20,B2为20,烟酸100,B6为20,泛酸50,叶酸5,B12为0.02,C为100,H为0.1,肌醇50,氯化胆碱250,添加量为0.1% 鲶鱼 微生素混合物组成(mg):每克预混料含B1为2,B2为3,泛酸钙6,尼克酸12,吡哆醇2,叶酸0.5,H为0.2,C为50,A为500IU,D3为25IU,E为20IU,K为0.5,氯化胆碱60,添加量0.4% (3)草食性鱼类复合添加剂配方? 草鱼?鲂鱼 矿物质或微量元素(mg):磷酸氢钙14415,硫酸亚铁250,硫酸锌200,硫酸锰92,硫酸铜20,碘化钾1.6,钼酸铵0.4,添加量为1.5% 微生素混合物组成(mg):每克预混料含B1为10,B2为10,B6为30,叶酸1,泛酸钙20,烟酸30,B12为0.01,C为10,K3为1,E为20,A为10000 IU,D为2000IU,添加量为0.1% 草鱼?鲂鱼 矿物质或微量元素(mg):每1千克含的克数硫酸镁150,食盐150,硫酸亚铁30,硫酸锌5,硫酸锰2,硫酸铜0.75,氯化钴0.08,碘化钾0.1,沸石粉662.1,添加量为1.5%~2% 微生素混合物组成(mg):B1为30,B2为30,B6为20,B5为5,A为50,D3为5.5,E为50,K3为25,烟酸100,泛酸钙50,叶酸7.5,玉米蛋白粉687,添加量为0.1% (二)基础饲料配方? 1?国外鲤鱼饲料配方? 成鱼: 饲料原料组成(%):蚕蛹粉30、米糠30、酱油粕20、大麦20 粗蛋白:30 饲料系数:3.3
罗非鱼介绍及图片 一、外貌特征 罗非鱼是一群中小型鱼类,它的外形、个体大小有点类似鲫鱼,鳍条多荆似鳜鱼。广盐性鱼类,海淡水中皆可生存。 罗非鱼养殖现在已经发展到全世界的许多国家,我国养殖的罗非鱼有莫桑比克罗非鱼、尼罗罗非鱼、奥利亚罗非鱼和红色罗非鱼4种。 二、生活习性 1、适应性强 罗非鱼生长与温度有密切关系,生长温度16~38℃,适温22~35℃,是一种广盐性鱼类,海淡水中皆可生存,耐低氧,一般栖息于水的下层,但随水温变化或鱼体大小改变栖息水层。 2、食性杂
罗非鱼是以植物为主的杂食性鱼类,池塘中的罗非鱼,消化道内含物大部分是有机碎屑及其他植物性饲料(如水草类、商品饲料等等),其 次是浮游植物、浮游动物和少量底栖动物。 3、耐低氧 罗非鱼耐低氧能力很强,窒息点为0.07~0.23毫克/升,水中溶氧1.6毫克/升时,罗非鱼仍能生活和繁殖。水中溶氧3毫克/升以上时, 生长不受影响。 4、不耐低高温 罗非鱼的生存温度范围为15~35℃。当水温低于15℃时,罗非鱼处于休眠状态。罗非鱼**临界温度约40~41℃,**适宜生长温度为28~32℃,罗非鱼繁殖温度在20℃以上。 5、繁殖能力强 罗非鱼性成熟早,产卵周期短,口腔孵育幼鱼,繁殖条件要求不高, 大水面积静止水体内自然繁殖。 三、营养价值
经测定每100克肉中含蛋白质20.5克,脂肪6.93克,热量148千卡,钙70毫克,钠50毫克,磷37毫克,铁1毫克,维生素B10.1毫克,维生素B20.12毫克。 1、罗非鱼蛋白质含量高,富含人体所需的多种必需氨基酸,其中谷氨酸和甘氨酸含量特别高,必需氨基酸组成平衡且含量丰富,属于优质高蛋白产品。鱼蛋白与肉类蛋白相比更易于人体消化和吸收,摄取的营养会被毫无浪费地充分利用。 2、罗非鱼视黄醇含量高,视黄醇有助于防治夜盲症、视力减退,有助于治疗多种眼疾,维护免疫系统正常功能,祛除老年斑,促进发育,强壮骨骼,维护皮肤、头发、牙齿、牙龈的健康,有抗呼吸系统感染作用。 3、罗非鱼牛磺酸含量高达210-220毫克/100克,牛磺酸可与EPA 和DHA媲美,能够有助于降低血液中胆固醇和中性脂肪,调节血压,强化肝功能,促进胰岛素分泌,促进视网膜发育,帮助视力恢复。而畜肉中牛磺酸的含量很低,如牛肉中含量仅为32毫克/100克,鸡肉中一点都没有。
鲤鱼饲料配方 (1)麸皮45%、豆饼40%、大麦10%、鱼粉5%,添加复合维生素、无机盐、赖氨酸、蛋氨酸适量,饲料系数为2,该配方由北京市水产科研所提供。 (2)麸皮45%、鱼粉30%、豆饼15%、大麦10%,添加剂量同(1),饵料系数为2,该配方由北京市水产科研所提供。 (3)豆饼50%、鱼粉15%、麸皮15%、米糠15%、复合维生素1%,无机盐、抗生素下脚料各1%、粘合剂2%,饵料系数为2.7,该配方由上海市水产科研所提供。 成鱼: 饲料原料组成(%):鱼粉10,虾糠20,麸皮50,玉米10,干薯粉10,维生素 0.3 粗蛋白(%):25.5 来源: 山东 、鲤鱼饲料配方: (1)贻贝粉15%,豆饼15%,麦麸45%,大麦10%,玉米15%。成鱼使用。 (2)贻贝粉5%,秘鲁鱼粉30%,豆饼15%,麦麸40%,玉米10%。成鱼使用。 (3)秘鲁鱼粉5%,豆饼40%,麦麸45%,大麦10%。成鱼使用。(4)贻贝粉5%,秘鲁鱼粉40%,豆饼15%,麦麸10%,大麦30%。鱼种使用。 (5)贻贝粉5%,秘鲁鱼粉45%,豆饼15%,麦麸10%,大麦20%,
啤酒酵母5%。鱼种使用。 2、草鱼饲料配方: (1)米糠40%,血粉12%,麦麸37.5%,豆饼10%,贝壳粉0.2%,尿素0.1%,生长素0.1%,食盐0.1%。 (2)稻草粉40%,大麦粉40%,菜籽饼15%,鱼粉5%,另加贝壳粉1%,食盐1% 3、罗非鱼饲料配方: (1)麦麸30%,豆饼35%,鱼粉15%,玉米5%,槐树叶5%,大8.5%,生长素1%,食盐0.5%。用于淡水养鱼。 (2)鱼粉10%,骨粉10%,豆饼35%,麦麸25%,干豆渣20%。 ?鲤鱼饲料配方 ? ?成鱼: 饲料原料组成(%):鱼粉12,豆粕25,花生粕10,菜粕13,玉米胚芽饼15,次粉麸皮20,添加剂5 粗蛋白:30 饲料系数:2.2 成鱼: 饲料原料组成(%):鱼粉12,豆粕25,菜粕20,棉粕7,玉米胚芽饼15,次粉麸皮16,添加剂5 粗蛋白:32.0 粗蛋白:30 饲料系数:2.2
罗非鱼养殖实用技术 罗非鱼(Tilapia)原产于非洲,是热水性鱼类,共有一百多种。我国先后从国外引进并已大量推广养殖的种类有:莫桑比克罗非鱼(俗称非洲鲫鱼)、尼罗罗非鱼以及奥利亚罗非鱼等。 第一部分罗非鱼的生物学特性 一、形态特征 属鲈形目,丽鱼科,罗非鱼体型侧扁,体披圆鳞,侧面线分上下两段,鳍较大,背鳍有15条以上的硬棘,软条8—12条,腹鳍硬棘1条,软条5条、臀鳍硬棘3条,软条9—11条,尾鳍后缘平截式略呈弧形,不分叉,体色因种类、环境及其生殖腺发育状况而有不同,有的体表和鳍上呈现黑色斑点或条纹,在繁殖期间体色变化较大。 尼罗罗非鱼的体型较大,背较厚、口较小。体长为头长的2.9—3.1倍,为体高的2.4—2.6倍。体色呈暗棕色,背部较深,自鳃盖后方至尾柄有时有8—9条黑色条纹。背鳍和臀鳍的软条膜有许多褐色斑点,尾鳍8—10条整齐的条纹,在生殖期间,雄鱼的胸鳍基部和尾端鳞片呈微红色,背鳍上缘红边不明显。 奥利亚罗非鱼体型基本与尼罗罗非鱼相似,但两者明显的区别是尾鳍的条纹状上,尼罗罗非鱼为黑色垂直条纹,奥利亚罗非鱼为紫色不垂直而为点状的条纹。 由于莫桑比克罗非鱼个体较小,已很少养殖,奥利亚罗非鱼较多的是作杂亲本使用,目前主要养殖品种是尼罗罗非鱼及通过杂交产生雄性化罗非鱼。 二、生活习性 罗非鱼要求较高的水温,适温范围是18—38℃,在26—30℃生长最快,低于15℃时行动呆滞,不摄食少动,据测定,尼罗罗非鱼的临界温度为8.61±0.15℃,而奥利亚罗非鱼为7.13±0.07℃。致死温度,尼罗罗非鱼为6.14±0.11℃,而奥利亚罗非鱼3.95±0.24℃。所以,在北方地区自然条件下饲养罗非鱼,应注意适时捕捞,对它的保种越冬工作不可忽视。罗非鱼能耐低氧,溶解氧降到1.5毫克/升时仍能正常生活,1.0—1.2毫克/升时出现浮头,0.3—0.4毫克/升时窒息死亡。 三、食性 罗非鱼是以植物性饲料为主的杂食性鱼类,在幼鱼阶段主要摄食游生物;成鱼的食物种类很多,各种藻类、嫩草、有机碎屑、底栖动物和水生昆虫等都是摄食对象,并能消化其它鱼类不能消化的蓝藻和绿藻,还能直接吞食各种人工饲料和部分有机肥料。
最全的钓罗非鱼资料 钓罗非杂谈(上) 一、罗非简介 罗非,又名非洲鲫鱼,为原产非洲内陆之热带慈鲷科,耐寒度较差,体型不大,经济价值极低。后经多次改良,不但成长迅速、繁殖力更强,且体形变肥硕、肉质转佳,耐寒度也降至摄氏8度左右,成为养殖主流鱼种之一。生长环境与水质良好的罗非鱼,没有土味或腥味,可媲美海洋鲷鱼,肉质细嫩且富弹性、骨少味鲜,可做生鱼片,在台湾被称为「台湾鲷」,已成出口大宗,加上跃居垂钓对象鱼之首选,是属低成本高利润的养殖鱼种。 罗非属底栖之广盐性鱼类,耐盐度与耐污染度颇高,食性虽广而杂,仍以偏好腥肉为主,食量颇大。群居性与领域性强,对它鱼种具攻击性,生性胆大粗犷,不像其它鱼种那么怕惊扰,无论声响、倒影,或直接扰乱水体,对它都不太有影响,甚至会跑到钓者脚边来觅食。因此,初学垂钩者,罗非鱼就是最好的垂钓对象。不过,罗非的腹鳍、背鳍坚硬刺手,被轻扎会血流不止、疼痛异常,甚至出现伤口愈合不易情形,所以摘鱼时要特别小心。唯其只在摄氏16~32度之适宜水温下,就饵意愿较高,所以垂钓罗非鱼,季节、温度与鱼层选择很重要,是一种「虽好钓、却不易钓好」的鱼种。 罗非遇缺氧时虽会浮头,但却属耐低氧的鱼种,公斤级大鱼甚至在离水二小时后仍可存活,显见其生命力之顽强。浮头后,常可见整群之罗非在水面张嘴,十分壮观;一个池子鱼量的多寡,从此处可一目了然,是无法掩饰的。当然,罗非既属底栖鱼种,此时就不用期待它们会就饵了,所谓「神仙难钓午时鱼」、「看得到、钓不到」,正是描述此一情境。虽整群浮头,不过,还是有一些方式可以让钓况逆转的,此是后话。 二、罗非的食性与口癖 全世界的罗非号称七百多种,名称之多可想而知。但一般人所说的罗非鱼,多指被养殖于人工池中,吃饲料长大,口癖变化不大的改良品种而言,体型硕大齐一,利于垂钓,常见偏名有福寿鱼(台湾)、尼罗鱼、非洲仔等。而那些流浪到野塘、水库或溪湖等水域繁殖的罗非鱼,品种则参差不齐,大小与口癖也不一致,甚至有只吃石上青苔者,很难一概而论,较不利于垂钓推广。 如果,有人问起「溪钓野罗非」该用何饵料时,千万不要回答「冷冻饵」,因为那些野罗非,多是识青苔却不知肉味为何物,未必会买帐。因习惯性的因素,罗非对完全没有尝过的饵料,需一段期间来适应,即使一个全新口味的高档「冷冻肉饵」,在青苔环伺的溪河中,也要给一点时间熟悉才行;等吃上瘾后,就会形成所谓的「口癖」,以后要不吃都很难了。 很多人钓罗非时常被饵料所困惑,面对眼花撩乱的偏方与添加物,心中不免存疑,但似
在黑坑钓罗非鱼饵料问题及解决妙策 罗非鱼,也叫非洲鲫鱼、福寿鱼,是一种择优引进的经济型鱼种,在南方水库、养殖塘较为常见,在温度较高的时候,罗非鱼也能适应,并且更加活跃,因此成为钓鱼人在夏天钓鱼时的主要目标鱼。下面小编整理了黑坑钓罗非的饵料配置问题和技巧经验,希望能给钓友们垂钓罗非鱼带来一些帮助。黑坑钓罗非疑问问:黑坑钓大罗非,窝里有鱼不开口,要怎么配饵料?问:黑坑罗非多半都是回锅鱼,一斤到三斤,坑里也有别的鱼种,小杂鱼不多,水深1,5米至2米5。平时都是冰冻饵加果酸蛋奶之类的,都钓不好,请问如何配饵料?高手回答窝里有鱼不开口或是鱼口不好,一般的情况下有以下五个原因。(1)饵料的味型不对,罗非是食肉鱼,对肝腥臭味情有独钟,因此,开料要腥、臭、香,相结合。如腥:冷冻饵,赤尾青,虾粉。臭:如黑水,王酱,乌鸡膏等。香:如鱼香宾,猪母乳,千里香等。 (2)子线和钩过大,适当换细一号的子线和钩。 (3)饵料过硬而挡口,可相应把饵料开得软一点,必要时可开昆布丝拉饵垂钓. (4)浮漂可能调得太钝,可适当调整。 (5)天气的原因,气压低,水缺氧,罗非上浮等也不吃钓。 总之,罗非越大就越精,吃饵也更谨慎,特别是回收回放和长期开钓的坑塘,由于不断的脱钩跑鱼,鱼对饵料有畏食性等,也会造成这种原因。以上谨供参考。各种小药的作用香虎:对钓饵有增香、甜等提味的作用,能增加饵料在水中扩散的范围,在寒冷、水脏等饵料味道不易扩散的水域施钓时,按照0.2%的比例添加,能提高钓效,添加过量会死窝,易招小鱼(皮肤病患者慎用)。蛋奶:添加香精,天气闷热时用酸味(酸蛋奶)效果更佳。虾粉:有安抚受惊罗非之功效,天气寒冷时需增倍添加,添加量可占到饵料的30%。综合以上,在我们掉罗非鱼时,这三样东西就可以琢量添加来提高钓效。猪母乳:鱼不开口时,水温低,刺激鱼的食欲,生口鱼很喜欢,但极易造成死窝,参考用量为每200克湿饵添加指甲的量。黑鸡膏:在避小鱼方面有一定的作用,添加后所上鱼体明显更大,建议用量150克添加花生米大小的量,野钓不建议使用,因为它的味道会永远留在水底土中,使此窝鱼成瘾成癖,极易造成偏食,其他人钓此窝就钓不好。 资深玩家的黑坑钓罗非鱼经验总结 熟悉黑坑钓的钓友们估计都遇到过钓罗非鱼时,鱼不吃饵的情况。虽然窝子里明显有鱼,可是它们就是不咬钩。只见浮漂在水里面像木桩一样,一动不动。那么遇到这种情况时,究竟该如何解决呢?其实说简单也简单,说难也难。首先钓者要保持头脑清醒,分析出现这种情况的具体原因是什么。通过这种仔细地观察和分析,采取具体的应对措施即可。一些钓者在遇到这种情况时,凭着以往的垂钓经验,想当然地换这个换那个,结果折腾了一番之后,仍是一点作用没有,浮漂还是一动不动,只是把自己搞得满头大汗,气喘吁吁。一些钓龄短的钓友甚至会
罗非鱼九种常见疾病的防治 一、肥胖症 [病原]长期投喂低蛋白、高脂肪、高糖类和缺维生素的饵料,造成罗非鱼脂肪代谢障碍,脂肪大量贮积,鱼体肥胖,抗病能力降低。 [症状与诊断]患肥胖症的罗非鱼呈全身性脂肪细胞增生、脂肪浸润,特别是腹腔内的脂肪组织以及脏器周围的脂肪组织显著增加,患病鱼腹腔内脂肪组织可达体重的3%-5%,肝脏呈淡黄色,肝组织高度脂肪变性,肝细胞萎缩。将整块肝组织剪下放在水中,会浮在水面上,而正常肝脏会立即沉入水底。 [流行与危害]工厂化养殖罗非鱼,密度高。若饵料营养不当,容易发生此病。该病主要危害性成熟个体。患病鱼抗病能力低,容易感染大肠杆菌和气单胸菌等病。肥胖症为条件致病菌的感染创造了条件。肥胖症继发细菌感染对罗非鱼造成更严重的危害。 [预防与治疗](1)改进饵料配方,尽量满足罗非鱼正常生长的需要。饵料中适当添加维生素B、维生素C和维生素E。也可增加一些天然饲料。(2)该病主要危害性成熟个体,应及时起捕成熟个体上市,可养活损失,缓解病情。(3)加强饲养管理,保持水质清洁水质清新鲜,定期服用抗菌素。 二、运动性气单胞菌病
[病原]为嗜水气单胞菌(Aeromonas hydrophila)。该菌广泛存在于正常鱼肠道中和池塘水中,属于条件致病菌. [症状与诊断]由嗜水气单胞菌引起的罗非鱼疾病,从症状表现来看,可以分为两个类型,即肠炎型和体表溃烂型.前者主要表现为肛门发红,肛门附近的皮肤发红;解剖观察可见肠道充血发红.后者主要表现为病鱼体表呈现斑块溃烂。 [流行与危害]肠炎型运动性气单胞菌病主要危害体重100克以下的罗非鱼,特别是10克以下的稚鱼.体表溃烂病在工厂化养殖和越冬期的罗非鱼鱼和和新鱼中均有发生.罗非鱼养殖密度高,容易造成鱼体损伤,加上溶氧低,水质条件差,病原菌数量增加快,很容易发生体表皮肤溃烂病.严重发病池感染率可在50%以上。 [预防与治疗] (1)用水体终浓度为0.5毫克/升-1毫克/升的呋喃唑铜全池遍洒,每天1次,连用2天.(2)1毫克/升漂白粉(含氯量30%)全池涮洒。(3)每100千鱼体重用2克呋喃唑酮拌入饲料投喂,每天1次,连用3天。(4)加强入期间的饲养管理。水温保持在20度,投饵应少而精,及时捞去残饵;注意经常换水,保持水质良好,定期洒些石灰水,使水保持弱碱性,可以预防该病的发生。 三、车轮虫病[病原]车轮虫。 [症状与诊断]车轮虫寄生于罗非鱼的皮肤,鳍和鳃等与水接触
3 国内外渔业发展概况 (2) 3.1世界渔业发展概况 (3) 3.1.1 世界渔业资源状况 (3) 3.1.2 水产品成为世界贸易大宗产品 (4) 3.1.3 世界水产品贸易概况4、5 (4) 3.1.4 世界水产品生产概况6、7、8 (5) 3.1.5 世界水产品加工业概况 (7) 3.1.6 世界先进渔业大国发展主要特色 (7) 3.1.7 世界渔业发展的趋势15、16、17 (11)
3 国内外渔业发展概况 3.1 世界渔业发展概况 3.2 我国渔业发展概况 3.3渔业产业结构的演化 3.4渔业经济增长方式的转变 3.5 渔业资源可持续利用与渔业可持续发展 3.6渔业发展的影响因素(资源、技术、资金、市场、政策、人才等)3.7 本章小结
3.1世界渔业发展概况 3.1.1 世界渔业资源状况 地球表面的总面积5.1亿km2,其中海洋面积为3.6亿km2,海洋占地球表面的总面积的71%。国际上把世界海洋划分为16大渔区,即太平洋和大西洋各分为西北、东北、中西、中东、西南、东南六部分,印度洋分为东、西两部分,地中海,黑海及南极海区。按照渔业资源的丰富程度,世界海洋分为四大渔场,即北太平洋渔场、东北大西洋渔场、西北大西洋渔场、和秘鲁渔场。 海洋蕴藏着十分丰富的海洋生物资源。世界海洋生物约20万种以上,其中海洋动物约18万种。从生物学上分,海洋生物资源包括鱼类资源、海洋无脊椎动物资源、海洋脊椎动物资源和海洋藻类资源。全世界鱼类约有2.5~3万种,其中海产鱼类超过1.6万种,但海洋捕捞种类的约有200种。其中年产量不足5万吨的占多数,约为140多种;超过100万吨的仅有12种,即狭鳕、大西洋鳕鱼、秘鲁鳀鱼、大西洋鲱鱼、鲐鱼、毛鳞鱼、远东拟沙丁鱼、沙瑙鱼、智利竹荚鱼、沙丁鱼、鲣、黄鳍金枪鱼等,它们约占世界海洋渔获量的1/3。海洋无脊椎动物估计有16万种,经济价值较大、目前已被人类利用的约有130多种,包括乌贼、章鱼、鱿鱼、贻贝、牡蛎、扇贝、蛤、蚶、砗磲、鲍鱼、红螺、对虾、龙虾、蟹、海参、海蜇等。大西洋西北部是世界上捕捞头足类的中心,年产约100万吨。大西洋中东部是世界上头足类捕捞的第二渔场,年产约30万吨。中国近海黄海、东海是以日本枪乌贼和大枪乌贼为主。据估计,世界大陆架和大陆斜坡上部海区内头足类的蕴藏量约800~1200万吨,有90%尚未开发。全世界有牡蛎200多种,中国沿海有20多种。贻贝有紫贻贝和翡翠贻贝、加州贻贝等。扇贝的种类也很多,分布广泛,世界各海洋都有。捕虾业是经济价值最高的一种渔业,世界上捕虾的国家达七、八十个,主要产虾国家是美国、印度、日本、墨西哥等。虾场主要分布在南美、中美、欧洲南部、中国、朝鲜和日本南部外海。蟹类种类很多,中国有600多种,绝大多数为海生,常见的有三疣梭子蟹、锯缘青蟹等。在世界上产量最多的是勘察加蟹和雪蟹,年产约15万吨。全世界的海参约1100多种,可供食用的约40种,从渤海湾、辽东半岛到北部湾的涠州岛、南沙群岛都出产海参。中国的海蜇资源是很丰富的,中国北方沿海常见的是海蜇、面蜇、沙蜇三种,分布于南海的是黄斑海蜇。联合国FAO对世界海洋渔业资源年可捕量总体估计是,经济鱼类1.04亿吨,经济甲壳类230万吨,头足类1000万至1亿吨,灯笼鱼类1亿吨,南极磷虾1亿吨以上。1、2 近几十年来,人类对海洋生物资源的过度利用和对海洋日趋严重的污染,使全球范围内的海洋生产力和海洋环境质量出现明显退化。 一是过度捕捞引起的海洋生物资源衰退。随着世界各国海洋捕捞能力日益增强,现代化的捕捞能力迅速提高,渔业资源因过度捕捞而逐渐衰退。大多数野生鱼种已被充分利用,越来越多的鱼种已经被过度捕捞。一些捕鱼行为,如底拖网捕捞回破坏海
非洲三大湖和三湖慈鲷简介 慈鲷原产于热带中南美洲、非洲及西印度群岛,因具有极高观赏价值而被引进全球各地,如今已是热带与亚热带地区最常见的外来鱼种。其中生活在非洲中部三大湖的慈鲷(非洲三大湖指坦干依喀湖、马拉威湖、维多利亚湖),因其变化无穷的色彩、强壮的体魄、优雅的体型、超强的水土适应能力,已成为观赏鱼种的最佳选择,这就是我们平时所熟知的三湖慈鲷。其实在非洲大陆慈鲷科鱼种被发现逾700余种,除了三湖慈鲷外还有众多的非三湖慈鲷,除了极西北和非南区域外,几乎随处可见其身影。 三湖慈鲷能够成为进化最为成功的鱼类之一,是因为它们有许多让其它鱼类无法企及的共性,可在非同寻常的环境中生存,比如生活在ph值高达8.5的水质中,盐水、温水、低溶氧水水域等,三湖慈鲷另一个种群庞大的原因是它独特的护卵方式,通过口腔孵育,这种独特的口卵方式,极大地保证了幼鱼的成活率。同时这也是三湖慈鲷进化成功的又一佐证。 三湖慈鲷中最引人入胜的当选坦干依喀湖慈鲷(简称坦湖慈鲷),因其在三湖中形成时间最久,湖深度也最深,各种地貌的冲击加上河流的流入,所以矿物质含量极高,湖水清澈,千变万化的体型、色彩和进食繁殖行为让爱好者更加投入其中。坦湖慈鲷既拥有体长90cm的全世界最大的90天使,也拥有像卷贝这种体长仅有3cm左右的袖珍慈鲷,多变的色泽虽然带来了区分上的困难,但相对也会给爱好者带来不同的乐趣。
相对于坦干依喀湖湖水的清澈透亮,马拉威湖的水体受地质关系的影响,沉淀物过多,不如坦湖清澈,而是有点浑浊,所以为了更加方面交配,马湖慈鲷一般以体色闻名于世,比如全身泛着金黄色光泽的非洲王子和拥有深蓝色光泽的阿里,可谓马湖慈鲷中最具代表性的鱼种。 维多利亚湖因人类环境的过度破坏,慈鲷种群已经大大减少,目前,维湖慈鲷大致分作五个属,虽然这些属在分类上还有不少争议,但是,还是可以就外型及食性来区分其差异性。维湖慈鲷比较著名的像黑金刚、绿魔王、翡翠王子、七彩天使等,色彩主要是由黑、红、蓝等色彩所组成。 前言:非洲是古老而神秘的,非洲东部的三大湖泊:坦干伊克湖、马拉威湖和维多利亚湖更是非洲慈鲷爱好者心目中的圣殿。 坦干伊克湖(Tangnyi Lake) 湖水澄明有如玻璃般的清澈,长650公里,深1500公尺,这个位于东非大地沟中的坦干伊克湖是世界上最老和第二深的美丽湖泊。 坦湖的美丽是大自然的杰作,而她的无尽资源则是大自然对人类的祝福,湖水中养育着超过300种的鱼类,比整个欧洲的鱼种还要多,其中不少的种类在全世界只有在这个湖看得到,光是和慈鲷有关的鱼种就占了湖中300多种鱼的三份之二。在它们这些鱼种之中“黄珍珠雀”总是会成群的游,每一只珍珠般色泽的鱼都在岩礁间狩寻小虾和其他微生物来进食;而另一种类的慈鲷
肉鸡饲料参考配方 1.肉雏鸡的饲料配方 (1)玉米55.3%,豆粕38%,磷酸氢钙1.4%石粉1%,食盐0.3%,食用油3%,预混料1%,“生物催肥精”+“粗饲料降解剂”(或“高效保健促长液”6斤)适量。 (2)玉米54.2%,豆粕34%,菜粕5%,磷酸氢钙.5%,石粉1%,食盐0.3%食用油3%预混料%,“生物催肥精”+“粗饲料降解剂”(或“高效保健促长液”6斤)适量。 (3)玉米55.2%,豆粕32%,鱼粉2%,菜粕4%,磷酸氢钙1.5%,石粉1%,食盐0.3%,食用油3%,预混料1%,“生物催肥精”+“粗饲料降解剂”(或“高效保健促长液”6斤适量。 (4)全价小鸡料100%(100市斤),“生物催肥精”+“粗饲料降解剂”(或“高效保健促长液”6斤)适量。 2.肉中鸡的饲料配方 (1)玉米58.2%,豆粕35%,磷酸氢钙1.4%,石粉1.1%,食盐0.3%,油3%,添加剂1%,“生物催肥精”+“粗饲料降解剂”(或“高效保健促长液”10斤)适量。 (2)玉米57.2%,豆粕31.5%,菜粕5%,磷酸氢钙1.3%,石粉1.2%,食盐0.3%,油2.5%,添加剂1%,“生物催肥精”+“粗饲料降解剂”(或“高效保健促长液”10斤)适量。 (3)玉米57.7%,豆粕27%,鱼粉2%,菜粕4%,棉粕3%,磷酸氢钙1.3%,石粉1.2%,食盐0.3%,油2.5%,添加剂1%,“生物催肥精”+“粗饲料降解剂”(或“高效保健促长液”10斤)适量。
(4)全价中鸡料100%(100市斤),“生物催肥精”+“粗饲料降解剂”(或“高效保健促长液”10斤)适量。 3.肉大鸡的饲料配方 (1)玉米60.2%,麦麸3%,豆粕30%,磷酸氢钙1.3%,石粉1.2%,食盐0.3%,油3%,添加剂1%,“生物催肥精”+“粗饲料降解剂”(或“高效保健促长液”12斤)适量。 (2)玉米59.2%,麦麸2%,豆粕22.5%,菜粕9.5%,磷酸氢钙1.3%,石粉1.2%,食盐0.3%,油3%,添加剂1%,“生物催肥精”+“粗饲料降解剂”(或“高效保健促长液”12斤)适量。 (3)玉米60.7%,豆粕21%,鱼粉2%,菜粕4.5%,棉粕5%,磷酸氢钙1.3%,石粉1.2%,食盐0.3%,油3%,添加剂1%,“生物催肥精”+“粗饲料降解剂”(或“高效保健促长液”12斤)适量。 (4)全价大鸡料100%(100斤),“生物催肥精”+“粗饲料降解剂”(或“高效保健促长液”12斤)适量。 将上述饲料混合均匀兑水(饲料相同重量)打堆覆盖1小时以上,即可饲喂。饲养诀窍:小鸡在出生三天左右时,按照每1000只鸡注射一盒“一针肥”(用蒸馏水、氯化钠注射液、葡萄糖注射液稀释200~500倍后注射),配合拌和“生物催肥精”+“粗饲料降解剂”(或“高效保健促长液”)的饲料,疾病极少、成活率极高,生长快,较常规养殖的鸡相比较,能提前7~15天出栏,鸡场臭味基本消除,效益显著提高,养殖轻松。典型示范养鸡技术请看后面内容。 肉鸭饲料参考配方 雏鸭(1~25日龄):玉米50%、菜籽饼20%、碎米10%、麸皮10%、鱼粉6.5%、骨粉1%、贝壳粉1%、食盐0.5%,添加剂1%,“生物催肥精”+“粗饲料降解
尼罗罗非鱼俗称非洲鲫鱼,属于鲈形目丽鱼科Cichlidae、罗非鱼属Tilapia(有称丽鲷科,丽鲷属)。学名Tilapia nilotica (linnaeus),原产于非洲约旦的坦噶尼喀湖,现已广泛为其他国家和地区所引进,是联合国推荐养殖的优质水产养殖品种。 一、形态特征 体侧扁,头中等大小,口端位;眼中等大小,略偏头部上方。成熟雄鱼颌部不扩大,下颌长为头长的29~37%。鳞大,圆形,侧线分上、下两段。上段侧线有鳞片18~24枚,下段侧线有鳞片12~22枚。沿侧线列鳞数30~35,通常32~33。背鳍发达,起点于鳃盖后缘相对,终止于尾柄前端;硬棘16~17,软条12~13;臀鳍末端超过尾柄,硬棘3,软条9~11;胸鳍较长,可达到或超过腹鳍末端,无硬刺,软条14~15;腹鳍胸位,硬刺1,软条15。尾鳍末端钝圆形。幼鱼尾鳍后缘平截,成鱼尾鳍后缘呈扇形。体色呈黄褐至黄棕色,从背部至腹部,由深逐渐变浅;喉、胸部白色。成体雄性呈红色;体侧有9条与体轴垂直的黑色带条,其中背鳍下方有7条,尾柄上有2条;背鳍边缘黑色,在背鳍和臀鳍上有较为规则的黑色斑纹;尾鳍和胸鳍的边缘红色,成体雄鱼显得特别鲜艳;雌鱼体色较暗淡,孵育期间呈茶褐色,体侧黑,体条纹特别明显,头部也出现若干不太规则的黑色条纹。 二、生活习性 尼罗罗非鱼是热带鱼类,适宜的温度范围16~38℃,最适生长水温24~32℃,在30℃时生长最快。致死温度上限为42℃,下限为10℃。15~14℃食欲减退。10℃完全不摄食。 尼罗罗非鱼耐低氧性较强,在水温22~25℃时,0.7mg/L溶氧,仅表现出微弱的浮头,但仍能摄食;在溶氧量为2.24mg/L时摄食旺盛。为保持正常生长,水体中溶氧量必须保持在3mg/L以上,低于0.1mg/L 窒息。氨氮1mg/L以下。pH值在7.5~8.5之间。二氧化碳在50mg/L以下。 该鱼属广盐性鱼类,能适应较大盐度范围的变化,可以从淡水中直接移入盐度为15‰的海水中,反之亦然。若从较低盐度(15‰以下)开始,逐步升高盐度,经短期驯化,最后能在30‰盐度的海水中正常生长,在40‰的盐度下仍能生存。 尼罗罗非鱼一般生活于水底层,随水温变化早晨游向中、上层,中午接近水表层游动,傍晚在中、下层活动,夜间与黎明静止于水底。幼鱼喜集群游泳,成鱼遇敌害或拉网时先跳跃后潜入水底软泥,露嘴于泥外而不动。 幼鱼期,几乎全部摄食浮游动物~轮虫卵、桡足类无节幼体和小型枝角类,随着个体的生长逐渐转为杂食性,其食物种类,在天然水体中,完全取决于水体中天然饵料的种类及数量,通常以浮游植物、浮游动物为主,也摄取栖底生物、水生昆虫及其幼虫,甚至小鱼、小虾,有时也吃水草等。成鱼期主要摄食浮游植物,其中蓝藻占70%,一些对于鲢、鳙等鱼类较难消化利用的藻类,该鱼都能较好的消化利用。该鱼对项圈藻的同化效率为75%、对微囊藻为70%,对鱼腥藻为75%,对菱形藻为79%,对小球藻为49%。在人工喂养的条件下,除摄食以上天然饵料外,还大量摄食各类商品饲料。如糠麸、油料饼粕、豆渣、酒糟等农副产品和食品加工副产品,以及人工配合饵料。利用各种商品饲料饲养该鱼,能获得很显著的效果,在生产中还可采取投饵与施肥相结合的方法,能取得较好的经济效益。 三、繁殖习性
收稿日期:2006210225;修订日期:2007206207 基金项目:国家重点基础研究发展计划(编号:2006C B101805)资助 作者简介:贝锦新(1977— ),男,广东揭西人;博士;主要从事鱼类分子免疫学研究。E 2mail :beijinxin @https://www.doczj.com/doc/762516471.html, 通讯作者:林浩然,E 2mail :lsslhr @https://www.doczj.com/doc/762516471.html, ;T el :020********* 综述 鱼类基因数据库与生物信息学在鱼类基因开发上的应用 贝锦新 张 勇 李文笙 刘晓春 林浩然 (中山大学水生经济动物研究所,广州 510275) APP LICATION OF GEN OME DATABASES AN D BIOINFOR MATICS IN EXP LORATION FOR FISH GENES BEI Jin 2X in ,ZH ANG Y ong ,LI W en 2Sheng ,LIU X iao 2Chun and LIN Hao 2Ran (Institute o f Aquatic Economic Animals ,Sun Yat 2Sen (Zhongshan )Univer sity ,Guangzhou 510275) 关键词:鱼类基因组;同线性分析;同源比对;基因开发 K ey w ords :Fish gen ome ;C onservation of synteny ;H om ology search ;Data mining 中图分类号:Q78 文献标识码:A 文章编号:100023207(2008)0320387206 近十年来,随着分子生物学技术的发展,各国际组织间的合作研究使得资源基因组计划得以普遍实施。各种动物、植物基因组数据库均在互联网上陆续发布,生物信息量随之迅猛增长,然而如何利用并发掘这些数据蕴涵的宝藏,从中提取解释生命个体生长发育、免疫调节以及病害控制等机理的宝贵信息,已成为人类所面临的巨大挑战。因此,以获取、加工、储存、分配、分析和解读生物信息为手段,综合运用数学、计算机科学和生物学工具的交叉学科———生物信息学[1]由此诞生。也正是依靠生物信息学的辅助,人类基因组计划最终在2003年基本完成,基因组序列数据的拼接和组装也陆续完善。 各种鱼类大约占据了50%的脊椎动物种类,有着长远的进化史 [2] 。因此,利用这些丰富的鱼类资源,研究与系统发育 和进化相关的种间保守或者特异的遗传和分子机制就显得相当重要。但相对于高等脊椎动物而言,鱼类的分子生物学研究比较滞后。在鱼类基因开发过程中,大部分是通过不同鱼类或者是其他高阶元物种的基因序列同源性,设计简并引物进行目的片段扩增;或者通过表达序列标签(Ex pressed se 2 quence tags ,EST )数据库发现并筛选鱼类功能基因。随着鱼类 的基因组全序列测序拼接工作的不断完善,通过生物信息学和分子生物学的方法,新的鱼类功能基因逐渐被发现,这种技术路线采用了反向生物学的原理,即从基因到蛋白质再到功能研究的途径来发现新的生物活性分子[3]。国外科学家已经筛选到一批与发育、生殖及免疫相关的功能基因,而国内对于鱼类新功能基因的开发研究起步较晚,因此,我们应当运用生 物信息学结合分子生物学的手段,发掘鱼类以及其他生物基因组数据库中的关键信息,开发并研究鱼类重要的功能基因,使得我国在这一领域能够有创新性的成果。为此,本文综述了目前鱼类基因组数据库的研究现状,并结合例子介绍利用这些数据库进行基因开发的几种方法。 1 鱼类基因公共数据库资源 111 斑马鱼(Danio rerio ,zebrafish ) 斑马鱼隶属鲤形目(Cypriniformes )鲤科,由于个体小、周期产卵且产卵周期短、产卵量高、卵大、体外受精、胚胎透明、体外发育、胚胎早期发育快和易于大量获得样品等独有的特点而成为目前世界公认的模式脊椎动物之一。单倍体斑马鱼基因组由25条染色体组成,含有117×103Mb (1Mb =106个碱基对或bp ),大约为哺乳类基因组大小的二分之一[4]。 2001年2月开始,英国Sanger 研究所开始了斑马鱼全基因组 测序工作;主要通过BAC (Bacterial artificial chrom os ome )、PAC (P12derived artificial chrom os ome )文库的克隆测序和拼接,以 及基因组鸟枪法测序(Whole genome shotgun sequencing )和组装两种策略。目前,该研究所和国际同行已经测序并拼接了 6653条DNA 片段,包含碱基约116×103Mb (参照:http ://w w w 1sanger 1ac 1uk/Projects/D 2rerio/)。许多不同的服务器都 提供了最新的斑马鱼基因组序列拼接数据,并能进行下载和比对(表1)。同时,以美国华盛顿大学为代表的许多研究所也进行了不同规模的EST 测序工作,这些丰富的斑马鱼EST 第32卷第3期 水生生物学报 V ol.32,N o.32008年5月 ACT A HY DROBI O LOG IC A SI NIC A May ,2008